Timing of hull split can vary depending on weather and variety. The almond industry’s leading variety, Nonpareil, typically splits in early to mid-July (photo by C. Parsons.)
Hull split in almond signals that it is time to apply treatments to control navel orangeworm. This timing is critical. Split hulls release a scent that attracts female NOW moths who lay their eggs on the suture of the splitting hull. Fungal spores can also invade the almond hull at split and cause hull rot.
In an Almond Board of California Training Tuesday webinar, almond experts recommended tree inspections starting at the top of the canopy on the southwest corner to detect onset of hull split.
Almond hulls split as the fruit ripens. If the entire suture on the hull opens and exposes the shell when the nut is squeezed, hull split has occurred. Timing of hull split can vary depending on weather and variety. The almond industry’s leading variety, Nonpareil, typically splits in early to mid-July.
It is the stage prior to a visible hull split that growers want to catch. When the hull is at the deep “V” stage, susceptibility to NOW infestations and pathogens that cause hull rot increase. Monitoring the crop for this stage is important.
Nuts mature faster at the top of the tree canopy, and that is where monitoring must begin. Using a ladder, lopping off upper branches of smaller trees or even using a pruning tower will allow close inspection of nuts from the top of the canopy to determine their stage of maturity.
Noting any blank nuts, or those lacking a kernel, is important as those are the first to initiate hull split, often several days before sound nuts. Mel Machado, vice president of grower services for Blue Diamond, calls blanks ‘a warning shot’ that alerts growers and farm managers that the hull split is imminent.
Experts advise paying attention to trees on the edge of the orchard as their nuts ripen earlier.
When planning hull split applications, they note that it is better to be early than late. Hull split applications should be made no later than at 1% hull split. That is the most effective time to spray for NOW as the timing often matches initiation of the second flight of NOW.
Machado also advises that once hull split begins, the entire orchard should be sprayed in five days or less. Since nuts at the tops of the trees are most vulnerable, consideration of aerial applications should be made to achieve best coverage. The best time to spray is early morning or at dusk.
Citrus leaf samples can be tested to positively identify Huanglongbing infection in a tree (photo courtesy S. Hajeri.)
Citrus Research Board reports that in addition to visual inspections, regular treatments and scouting for Asian Citrus Psyllids, growers are conducting routine testing of leaf samples to positively identify Huanglongbing (HLB) infection in a tree. The testing involves direct detection of the bacteria that causes HLB.
CDFA has provided leaf collection and handling protocols. They include a visual assessment of each tree to be sampled and looking for known HLB symptoms. Searching for and collecting symptomatic leaves when possible is recommended. If yellow shoot symptoms are present, 12 leaves from that branch should be collected. The petiole must be attached to the selected leaves. If yellow shoot symptoms are not present, the entire tree should be inspected for other HLB symptoms, including leaf mottling, twisted leaf ACP damage and vein thickening.
Next, the tree should be divided into four quadrants (north, south, east and west), and four leaves should be collected from each quadrant for one sample. Leaves selected should be young and of medium size, one growth period old and as near to flush as possible.
If no symptoms are present in the tree, select 16 leaves, four from each quadrant, from fully expanded current season flush. Wipe or brush leaves to remove dust and debris and check each sample to ensure there are no thorns and all insects and their life stages are removed.
Next, fold each sample at the mid rib. Wrap the folded bundle of leaves in a dry paper towel and place the sample in a zip-lock plastic bag. The bag should be labeled with the unique sample identifier number, the date collected and the exact location of the host tree, including address, cross street, city and county. The labeled bag should be placed inside another zip-lock bag and placed in an ice chest with blue ice packs to keep the samples cool.
CDFA notes that it is essential to place protective material between the ice packs and samples to ensure they stay dry and do not get freezer burn.
The agency also emphasizes making sure no ACP are with the leaf samples or inside the package.
Growers or orchard managers must mark the trees from which the samples came from for retesting purposes. If any samples test positive for the bacteria, CDFA will be notified and will contact the grower to resample.
Currently, the Citrus Pest Detection Program (CPDP), operated by the Central California Tristeza Eradication Agency, is permitted to provide HLB testing services of plant samples from growers throughout the state, except in HLB quarantine areas, via mail. However, growers located in the San Joaquin Valley can request CPDP representatives take samples from their orchard for testing rather than mailing.
Samples can be sent to the Citrus Pest Detection Program at Attn: PCR Laboratory 22847 Road 140 Tulare, CA 93274. The Tulare laboratory director, Subhas Hajeri, would like to speak with growers or PCAs before they bring samples to the lab. The phone number is (559) 686-4973.
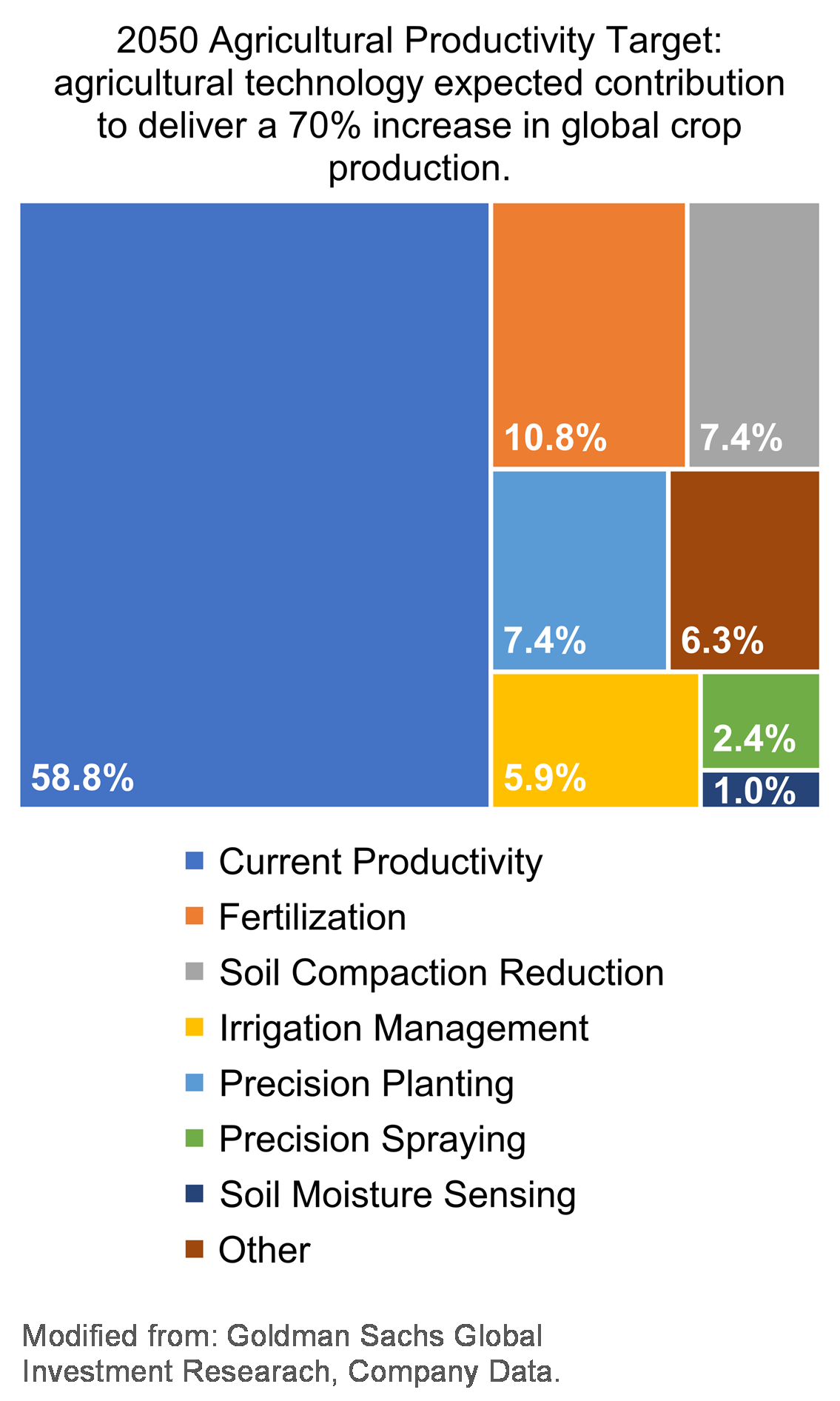
The global population is projected by the United Nations to reach 9.7 billion people by 2050. To satisfy the needs for food and fiber for that many people, agricultural production should increase by >70%. Globally and here in the U.S., studies suggest that improved management of soil resources, irrigation water and nutrients through digital agriculture (See Sidebar Digital Agriculture on page 9.) tools can be the key to increasing agricultural production (Fig. 1). According to USDA, sustaining the economic viability of agriculture is only one of the goals that must be achieved to fully satisfy the long-term food and fiber needs of the American people. Other key goals are to reduce the environmental footprint of agriculture and to improve the quality of life for farm families and communities.
Figure 1: Economic feasibility analysis showing the relative value that agricultural technologies are expected to add to the current global crop production to achieve the United Nation’s 2050 food production goals. Data extracted from Goldman Sachs, July 2016, “Precision Farming: Cheating Malthus with Digital Agriculture”
Improving Production Systems
Agriculture in the Southwestern U.S. is vital to the prosperity of rural communities and the national economy, both for internal consumption and for international export. Many crops are grown in the Southwestern U.S., including field crops and many specialty fruits, vegetables and nuts. In particular, around $12 billion/year in income is generated by agriculture in California’s Salinas River Valley and in the Colorado River Basin, including regions in Southern California that use Colorado River water for irrigation. In these regions, agriculture employs more than 500,000 workers.
Sustaining and improving the agricultural productivity in the region in the long term is, however, under threat as growers face major challenges: climate change, disease and pest outbreaks, increasing salinity and diminished and/or degraded soil and water resources, to mention a few. The region has experienced prolonged and major droughts since 2000, with the possibility of streamflow reductions by more than 50% by 2100. Future requirements to preserve groundwater may remove 500,000 acres from production in California alone. Minimizing salt and nutrient loading is increasingly mandated for effective water reuse. Uncertainty in the water supply in these regions is particularly troublesome, as agriculture there consumes 39% of the U.S. total irrigation water (almost 32 million acre-feet per year.) Additionally, climate change threatens to increase insect, pathogen and weed pressures and geographic distribution, while public interest and increasing on-farm costs push toward reducing pesticide use and organic farming.
Our current understanding of agricultural production systems indicates that Crop Yield is a complex function of Genetics × Environment × Management × Space × Time interactions. Understanding why yield deviates from optimal over space and time in different landscapes is key to adjusting management cost-effectively. When looking at the multi-year productivity of farmland, a rule of thumb might say that yield varies over time in response to climate as much as it changes across different spatial scales (within a field and across multiple fields) due to the variability in soil and landscape features.
Failure to adapt management to the dynamic spatial and temporal variability of crop growth often results in crop loss or over-application of agronomic inputs, which can lead to economic loss and environmental degradation. Real-time crop growth models that can leverage information from very high spatial and temporal resolution satellite imagery and ground networks of sensors, such as weather stations, are great candidates for guiding site-specific tailored agronomic management. Moreover, when combined with artificial intelligence, crop growth models can help improve farm management while considering the tradeoffs between different sustainability aspects at different spatial scales. For example, how to increase field scale profitability while reducing regional-scale environmental impacts.
Current Research
With the overarching goal of increasing agricultural profitability by reducing and optimizing inputs to increase yield and curb losses from abiotic and biotic stressors in the Southwestern U.S., UC Riverside recently started a five-year project on the use of artificial intelligence and big data from high-resolution imagery and ground sensor networks to improve the management of irrigation, fertilization and soil salinity as well as to enable early detection of weeds and pests. The project is led by Elia Scudiero, a professional researcher in UC Riverside’s Department of Environmental Sciences, and includes several co-investigators at UC Riverside and UC ANR, USDA-ARS, University of Arizona, Duke University, Kansas State University and University of Georgia. To accomplish its goal, the project relies on many collaborations with ag tech industry partners, including Planet Labs, Inc. Through the collaboration with Planet Labs, the project investigators will use daily high-resolution (around 12 feet) satellite imagery to monitor crop growth and soil properties.
Some of the artificial intelligence applications that the project will develop, for a variety of crops, include crop inventorying, soil mapping, estimations of crop water use and requirement, estimation of plant and soil nutrient status, analyzing the feasibility and cost-effectiveness of variable-rate fertigation across selected regions in the Southwestern U.S. and detecting weeds and pathogens. In the remainder of this article, we will provide a general overview and some preliminary results from some selected applications: Water use and water requirement estimations, mapping soil salinity with remote sensing and detecting biotic stressors.
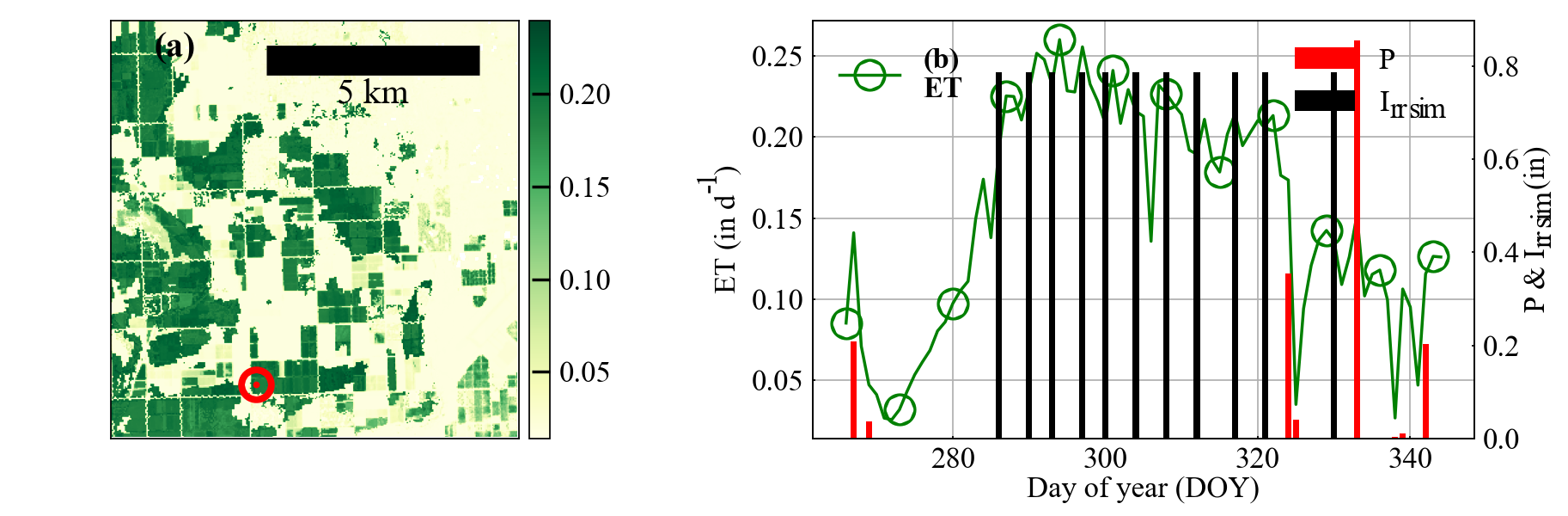
Figure 2: Left panel shows the map of evapotranspiration on November 10, 2019 for a selected region near Yuma, Ariz. using Landsat 8 imagery. This map has a red circle that is shown in the panel on the right illustrating evapotranspiration (ET) for the selected point for the season. Vegetation data from Landsat are combined with high-resolution meteorological data to estimate ET on an hourly time basis. The red bars illustrate rain events and the black bar illustrates the amount and timing of irrigation that would be needed to avoid unacceptable soil moisture depletion.
Water Use and Water Requirement Estimations
A key element of this project will be to provide reliable estimates of crop water use and irrigation forecasting at a very high resolution daily. Current remote-sensing-based evapotranspiration models often suffer from infrequent satellite overpasses, which are generally available weekly or every two weeks at the 30- to 100-foot spatial resolution. Such sporadic information is a limitation, especially for vegetable crops, which can have very fast-growing cycles. To overcome this and other limitations, the project is integrating daily meteorological information (from state and federal networks) and high-resolution satellite data from Planet Labs with BAITSSS, an evapotranspiration (ET) computer model developed by Dr. Ramesh Dhungel (a Research Scientist in the project) and colleagues. The current version of BAITSSS leverages information from the Landsat 8 satellite platform (NASA). An example application of the model is shown in Figure 1, where BAITSSS is combined with soil moisture modeling to forecast when irrigation is needed to supply crop water demand.
The project’s lead on water use and requirement estimations, Dr. Ray Anderson (USDA-ARS U.S. Salinity Laboratory, Riverside, Calif.), says that “along with irrigation management, we believe the daily imagery from Planet Labs will allow growers and irrigation managers to see evapotranspiration anomalies within a field. These anomalies could indicate irrigation issues or decreases in plant health due to other abiotic stressors such as salinity or nutrient deficiency. Identification of these anomalies will ensure more efficient field scouting and earlier identification of issues before permanent yield loss occurs.”
Mapping Soil Properties
Accurate knowledge of spatial variability of soil properties, such as texture, hydraulic properties and salinity, is important to best understand the reasons of crop yield spatial variability. Scudiero’s Digital Agronomy Lab at UC Riverside is developing novel tools based on machine learning to automate near-ground sensing of soil properties and remote sensing of soil salinity in irrigated farmland. Soil salinity maps are very useful to inform field-scale irrigation practices (e.g., to calculate the amount of irrigation water needed to avoid harmful accumulation of salts in the soil profile.)
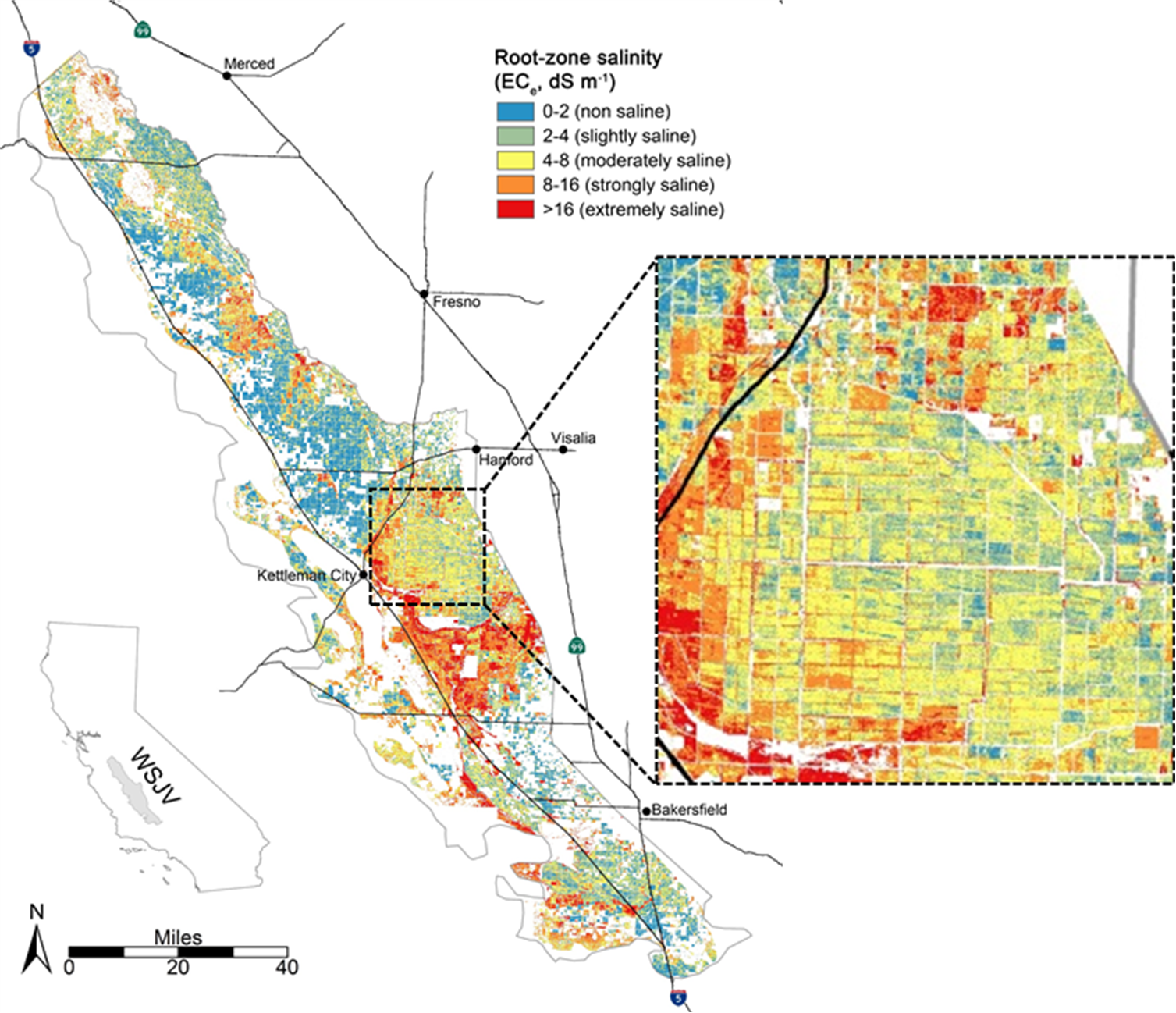
In this project, Scudiero and his team will be using field-scale soil maps of soil salinity collected in the past (since the early 1980s) and throughout the next five years by UC Riverside, the USDA-ARS U.S. Salinity Laboratory, the University of Arizona and other collaborators to generate soil salinity maps for the entire Southwestern U.S. Figure 3 shows the soil salinity map produced for the western San Joaquin Valley by Scudiero and colleagues (see the additional resources section.) In particular, Scudiero’s team will use the ground information to calibrate the Planet Labs time-series imagery to predict soil salinity in the root zone (e.g., the top four feet of the soil profile.) Satellite imagery alone is generally not sufficient to predict soil salinity. Other stressors (water stress, nutrient deficiency) have similar imagery properties to salinity. However, in short periods (two to five years), salinity remains fairly stable throughout the soil profile, contrary to other more transient stressors. Because of these differences in temporal variability between stressors, multi-year time series can be used to detect and map crop health reduction due to soil salinity.
Figure 3: 2013 remote sensing soil salinity (electrical conductivity of a saturated paste extract, ECe) for the zero to four feet soil profile in the western San Joaquin Valley. The map was generated using Landsat imagery with a resolution of about 900×900 ft. Current research is investigating the use of Planet imagery with a resolution of 12×12 ft to generate soil salinity maps across all irrigated farmland in the Southwestern U.S.
Biotic Stress Detection
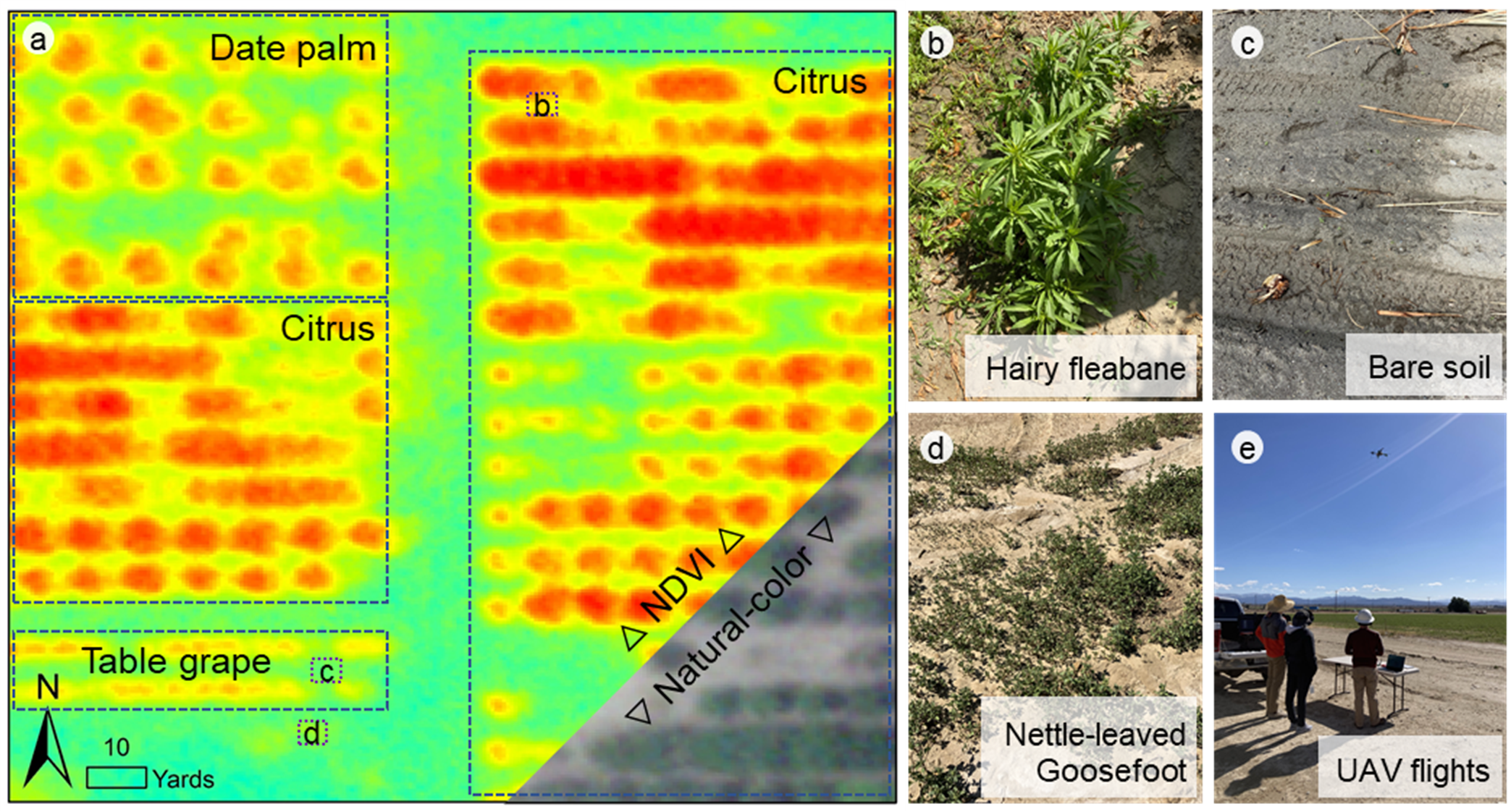
Within this project, UC Riverside scientists are using smartphone pictures, drone imagery and Planet Labs imagery (20 inch and 12 feet resolution) to develop artificial intelligence classifiers for early biotic stress detection. Figure 4 shows an example of a survey carried out by project collaborators Sonia Rios (UC ANR) and Robert Krueger (USDA-ARS) at UC Riverside’s Coachella Valley Agricultural Research Station in Thermal, Calif. Preliminary analyses show that 20×20-inch resolution imagery from the Planet SkySat satellites can be used to identify the presence of weeds against bare soil ground coverage in citrus and date palm. Single-date imagery cannot be used to successfully distinguish between different weed species. The project investigators hope that repeated satellite imagery, together with drone imagery, will be successful at identifying the emergence and extent of weeds and other biotic stress at the field and farm scales. To develop such tools, project investigators are carrying out controlled experiments at the UC Riverside research farms with controlled weed and pathogen pressures on a variety of vegetable crops.
Figure 4: Planet SkySat imagery from UC Riverside’s Coachella Valley Agricultural Research Station. In panel A, the high-resolution (~20×20-inch resolution) is shown in natural colors and using the Normalized Difference Vegetation Index (NDVI). The different crops present at the site are highlighted in panel A. Panels B, C and D show pictures taken as a ground-truth for the machine-learning weed classifier under development in the project. The project is also collecting imagery from unmanned aerial vehicles (UAV) at selected test sites to detect biotic stress (weeds and pathogens), panel E (photos courtesy R. Krueger (B, C and D) and E. Scudiero (E).)
Training to Growers and Consultants
From conversations with stakeholders in California and other states in the Southwestern U.S., we learned that most growers are committed to maintaining the quality and profitability of soil, water and other natural resources in the long term. Additionally, many growers recognize the potential of integrating digital agriculture technologies in their daily decision-making. Nevertheless, investing in new technology always requires considerable commitment. Therefore, growers would like to have more information on the cost-effectiveness of state-of-the-art technologies and on the suitability of technology to their local agricultural system. To address these and other questions, the project team is establishing a multi-state cooperative extension network to develop training programs for growers and consultants on the topics of precision agriculture, digital agriculture and the use of soil and plant sensors in agriculture. These training activities, scheduled to start in 2021, will include contributions from university personnel and industry members.
This article provided an overview and some preliminary results for the University of California Riverside, or UCR, -led project on “Artificial Intelligence for Sustainable Water, Nutrient, Salinity, and Pest Management in the Western US”. The project is funded by U.S. Department of Agriculture’s National Institute of Food and Agriculture (Grant Number: 2020-69012-31914). Through September 2025, the project will investigate the use of daily high-resolution satellite imagery and data science to identify inefficiencies in agronomic management practices and to support improved irrigation, fertilization and pest control in the irrigated farmland across the Colorado River Basin and Central and Southern California.
Additional information on the project and the content of this article can be requested from Elia Scudiero (elia.scudiero@ucr.edu). Information about the project’s cooperative extension events can be requested from ai4sa@ucr.edu. Further information about research on remote sensing of soil salinity and evapotranspiration can be found in the additional resources section.
Additional Resources: Dhungel, R., R.G. Allen, R. Trezza, and C.W. Robison. 2016. Evapotranspiration between satellite overpasses: methodology and case study in agricultural dominant semi-arid areas: Time integration of evapotranspiration. Meteorol. Appl. 23(4): 714–730. doi: 10.1002/met.1596. Scudiero, E., Corwin, D.L., Anderson, R.G., Yemoto, K., Clary, W., Wang, Z.L., Skaggs, T.H., 2017. Remote sensing is a viable tool for mapping soil salinity in agricultural lands. California Agriculture 71, 231-238. doi: 10.3733/ca.2017a0009
Sugarcane aphid on a forage sorghum leaf. Threshold for treatment is 50 aphids per leaf (photo by Pete Goodell, UC IPM Advisor Emeritus.)
Scouting forage sorghum fields early in the growing season for sugarcane aphid infestations may save considerable crop damage.
UCCE Farm Advisor Nick Clark, in a presentation for the virtual UC Alfalfa and Forage Field Day, said this invasive crop pest reproduces rapidly, and infestations have the potential to cause severe crop damage if not recognized and controlled early.
Dairy producers and those who grow forage crops for dairy silage plant forage sorghum for its drought tolerance and high yields. In 2016, the sugarcane aphid, an invasive pest from Mexico, spread into California forage sorghum fields and caused crop losses.
Sugarcane aphids suck plant juices and excrete honeydew. Their feeding on forage sorghum causes stunting, delayed development, diminished grain production and early senescence. The honeydew causes sooty mold on plants, affecting photosynthesis.
Field trials have shown that early infestation by this pest causes higher yield loss. In 20% of infested fields with no treatment, crop loss of 80% to 100% can be sustained if infestation occurs in the pre-boot stage. Infestation in the boot stage can cause yield loss of 52% to 69%. At panicle emergence, yield loss can be 67%.
Sugarcane aphids can be light green to orange in color. They are distinguished by black antennae, black cornicles and black feet.
Sugarcane aphid infestations are easy to spot, Clark said. Honeydew makes the leaves appear shiny and aphids are easily visible on the leaves. He advised scouting field edges weekly until sugarcane aphid is identified in the field. When sampling, he said to take leaves from plants 50 feet in from the field edge. Pull top and bottom leaves from 15 to 20 plants. The treatment threshold is reached when 25% of the samples have 50 or more aphids per leaf. 50 aphids can cover a dime-size area on the leaf.
Preventative controls for sugarcane aphid include managing Johnsongrass, which can serve as an overwintering host to sugarcane aphid, planting the forage sorghum crop as early as possible, keeping plants healthy and vigorous and using neonicotinoid-treated seed.
If chemical control is necessary, an aphid-specific treatment recommendation is flupyradifurone (Sivanto) in a water volume high enough for good foliar coverage.
Clark noted that growing populations of natural enemies including parasitoid wasps have been observed to control sugarcane aphid, but not always prevent crop damage.
In winegrape production, wildfire smoke reduces the amount of sunlight reaching the vines, increasing their stress level and affecting fruit quality. Smoke taint can alter flavor of grapes (photo by Jack Kelly Clark, UC IPM.)
Crop S.A.F.E. is a new tool developed to help wine grape growers manage risk from smoke taint due to wild fires and help protect workers.
Wildfires are an annual threat to fruit, vegetable and other crops grown where wind, high temperatures and smoke are present.
Smoke And Fire Events (S.A.F.E.) platform provides remote intelligence to enable informed crop management decisions during wildfire season. UPL, in collaboration with 6th Grain Corporations, a digital agriculture technology company, is introducing Crop S.A.F.E. as an online application for growers and crop managers.
In grape production, wildfire smoke reduces the amount of sunlight reaching the vines, increasing the stress level and affecting fruit quality. Smoke taint can alter flavor of grapes.
Molly E. Brown, chief science officer with 6th Grain, explains that Crop S.A.F.E. provides growers and managers with more in-depth information about smoke than what they might obtain from weather reports. The information can allow them to determine management for vineyards and reduce concentration of smoke-related aromas, flavors and compounds in the final wine product.
Examples of management strategies to prevent smoke taint, Brown explained, include washing canopy leaves after a smoke event to remove ash. A grower or manager could also choose to hand-harvest a vineyard to minimize skin breakage or rupturing for as long as possible or to harvest earlier to reduce smoke exposure.
The system can also provide regional information about smoke impacts in other areas and states. Brown said knowing how many days a particular vineyard or area has been exposed to smoke throughout the year is a critical part of determining wine quality in the coming year.
In addition, this tool helps keep workers as well as managers in the field safe from smoke exposure and breathing difficulties.
Crop S.A.F.E. uses Aerosol Optical Depth from NOAA to estimate the intensity of smoke exposure accumulated over time. The satellite-sourced map shows regions with high accumulated risk of smoke taint for grapes and other produce as well as field activities. The online resource estimates the intensity of smoke and other atmospheric hazards such as ozone and particulate matter levels plus temperature history.
Brown reports that merging information on field management with satellite observations of weather, crop extent and crop health allows growers and managers to plan to avoid negative impacts of smoke taint.
“By knowing the level of exposure to smoke and avoiding working in high smoke areas, workers can reduce risk of breathing difficulties,” Brown said.
Field trial at the UC Desert Research and Extension Center studied water and nitrogen efficiency in carrot production, comparing sprinkler and furrow irrigation and managing N applications (photo by A. Montazar.)
Trials conducted at the UC Desert Research and Extension Center and several commercial fields in Holtville are focused on best irrigation and nitrogen management in carrots. One of the major commodities grown in Imperial County, carrots are grown on an average of 16,000 acres.
Ali Montazar, UCCE irrigation and water management advisor in Imperial, Riverside and San Diego counties, said nitrogen and irrigation management in carrot production is critical for increasing efficiency in crop production and decreasing costs and nitrate leaching.
In desert carrot production, a common irrigation practice in stand establishment is to sprinkler irrigate the field every other day during the first two to three weeks after seeding. Many fields are furrow irrigated after germination. Montazar said a comparison of applied water and crop water consumption indicates that carrot fields could be over-irrigated during the stand establishment period.
There is a wide range in the length of carrot crop season anywhere from 125 days to 190 days. Montazar said the trials showed that carrot fields may also have variable water requirements due to soil types, early or late planting dates and length of crop season. A peak daily crop water use of 0.21 inches on March 23, 2020 151 days after planting was observed in a fresh market carrot field.
Use of sprinkler irrigation during the critical fruit set period allows for more frequent and lighter water applications than furrow irrigation and helps avoid wet and dry soil cycles which contribute to splitting of roots. Sprinklers also reduce salinity.
Montazar said that given an irrigation system efficiency of 70%, the approximate gross irrigation for carrots in that area is 1.9 to 2.1 acre feet, not including pre-irrigation.
Preliminary results of the trials show a notable amount of N uptake both in the roots and tops at harvest. Nearly 50% of the seasonal N accumulated in the tops and roots at 85 to 90 days after planting when the canopy is fully developed. An effective N application could be splitting N into 10% to 15% at planting, and the remainder through irrigation over the season, completed 15 to 20 days prior to harvest.
Montazar noted that 45% to 55% of total N accumulated in the carrot plants, mainly tops, are left in the field as residual soil N and can be a source of N for the next season.
This project was funded by the CDFA FREP and California Fresh Carrot Advisory Board.
Horseweed infesting a young pistachio orchard can steal water and nutrients from trees and interfere with irrigation efficiency (photo by C. Parsons.)
Weed control in tree nut orchards can be a moving target. New species can be introduced, herbicide application timing varies depending on the weed type and weed species can become resistant.
Many management tools are available for growers and farm managers, but methods of use can vary from year to year and from orchard to orchard.
Weeds affect young orchard growth by competing for water and nutrients. Large weeds can provide habitat for vertebrate pests, insect pests and contribute to the presence of nematodes and disease.
Almond Board of California’s Senior Specialist in Pest Management Drew Wolter said changes in weed populations from year to year, plus new, invasive weed species, challenge tree nut growers every year.
Weather conditions can influence weed populations. Salt or drought tolerance in a weed species allows it to proliferate in those conditions.
Soil texture and organic matter also affect weed populations. Annuals, including Puncture vine and horseweed, and perennials, such as Johnson grass, nutsedge and Bermuda grass, can dominate in lighter textured soils. Heavier soils will host curly dock, field bindweed and dallisgrass.
The UC IPM guidelines report that higher rates of preemergent herbicides may be necessary in clay or clay loam soils to achieve the same level of weed control as a lower rate in lighter soils.
Soil texture and irrigation methods also play a role in weed germination and establishment.
Horseweed, Italian ryegrass and field bindweed are three major weed species that invade tree nut orchards. Palmer amaranth and another amaranth species, such as hempweed, are two newer weeds that can pose challenges to control due to herbicide resistance. This is particularly problematic since there are few products registered for managing weeds in young orchards.
Horseweed’s prolific seed production makes this weed difficult to control. Herbicide control is more effective when this weed is in the rosette stage.
Field bindweed is one of the most difficult weeds to control, Wolter said. Its persistence is due to a vigorous horizontal rhizomatous system that can give rise to new roots and shoots once aboveground vegetative matter is knocked down or when roots are disturbed.
Systemic herbicides, such as glyphosate, should be applied when the plant is in its full bloom stage. During this stage of growth, the roots’ carbohydrate reserves are at their lowest, and the sugars produced in the leaves are being moved downward to the root system.
Italian ryegrass’ genetic diversity makes it difficult to develop a specific management plan. A winter annual, its seeds can germinate year-round in some conditions.
Globally, invasive insect pests cause substantial damage to agricultural crops and natural environments. In the U.S. alone, crop and forest production losses from invasive insects and pathogens have been estimated at almost $40 billion per year (Pimentel et al. 2005). On average, a new non-native invertebrate is introduced into California every 40 days. With respect to insects and mites, about one-third of these become pests (Dowell et al. 2016), and economic losses in California are estimated at more than $3 billion per year (Metcalf 1995).
Rapid growth of U.S. demand for fresh avocados has increased the fruit’s prominence in retail sales and diets. California is the largest producer of avocados grown in the U.S. The value of U.S. avocado production measured at approximately $392 million in 2017, (NASS, 2018). There are more than 3,000 avocado growers in the state farming on approximately 50,000 acres of land, with Ventura County leading the state in most acres planted and harvested in recent years (California Avocado Commission [CAC], 2020).
California avocados are particularly special because less than 1% of the state is suitable for growing them. A healthy, single ‘Hass’ avocado tree can produce up to 200 pounds of fresh fruit each year, which is ~500 pieces. In 2018 alone, California produced around 350 million pounds. Avocados are considered a specialty crop and are not cheap to grow because of water and land costs, and fruit therefore usually demands a high premium price.
Traditionally, insecticide use in California avocado orchards has been minimal due to relatively few pest species, which, for the most part, have been under adequate levels of biological control (Hoddle 2005). However, due to the increase of international trade, illegal importation and the smuggling of foliage, branches with leaves, whole plants and budwood, invasive insect and mite pests have begun to threaten the economic viability of avocado production in California.
Beetle gallery formation in an avocado branch (photo by A. Eskalen.)
Polyphagous and Kuroshio Shot Hole Borers
Polyphagous Shot Hole Bore (PSHB) is an invasive ambrosia beetle from Southeast Asia that was first detected in Los Angeles County in 2003 (Gomez et al. 2018, Rabaglia et al. 2006). PSHB is a pest of great concern in Southern California as it can attack over 300 tree species and reproduce on a subset of these that are capable of supporting beetle reproduction and growing the fungi that cause fusarium dieback.
PSHB has a strong symbiotic relationship with several fungi, including Fusarium euwallaceae and Fusarium kuroshium, the causal agent of Fusarium Dieback (FD) disease (Eskalen et al. 2013). The list of reproductive hosts includes several native oaks, maples, sycamores, willows and avocado. The fungus disrupts the vascular transport of water and nutrients on their host tree that eventually causes branch dieback. The most common symptoms of the disease include sugar or gum exudates, dieback, wilt and ultimately host tree mortality. Reports of dieback symptoms in El Cajon, San Diego County led to the discovery of another species, the Kuroshio shot hole borer (KSHB) in 2013.
KSHB is also present in commercial avocado orchards in San Diego County. KSHB was originally limited to the San Diego region but has since spread to Orange, Los Angeles, Ventura and Santa Barbara counties. Presumably, PSHB and KSHB were introduced accidentally into Southern California via wooden products and/or shipping material (e.g., pallets and dunnage) from Southeast Asia. KSHB is morphologically indistinguishable from PSHB, but species can be separated using molecular techniques.
The fungus gets into the woody tissue of the tree when a female PSHB excavates a gallery and inoculates the gallery walls with fungal spores that it carries in special structures called mycangia (Beaver, 1989). The female PSHB and her offspring feed exclusively on the fungal spores. PSHB adults engage in sibling mating within the natal gallery. Offspring sex ratios are female biased to maximize reproductive output such that one male can inseminate many of his female siblings. Once fully developed and mated, female PSHB exit their natal gallery to start a gallery of their own and repeat the cycle. Unlike their sisters, male PSHB do not disperse since they are not capable of flying and do not possess mycangia, which are specialized structures for fungus storage.
Pesticide efficacy may be limited, and this is attributed to the fact that PSHB feeds on fungi and not directly on wood tissue. This reduces exposure to active ingredients when feeding (Eatough Jones et al. 2017). Cultural management practices include: 1) removal of infected branches to reduce local beetle numbers and fungal spread in the infested area; 2) chipping (< 1 inch) and solarizing infested wood; and 3) avoiding movement of infested materials to new areas (if transportation of materials is necessary, it must be covered(UC IPM 2017).
These management strategies can perhaps abate the severity of local infestations. Also, limiting the spread of infected/infested trees include minimizing firewood transportation. Sterilizing pruning tools with household bleach or another cleaning solution can reduce the spread of the FD pathogen. The PSHB was found in Israel in 2009 in commercial avocado orchards where it damages trees. To date, there has been no avocado tree fatality due to the PSHB or FD in California. For more information regarding PSHB, visit ucanr.edu/sites/pshb/.
Severe Laurel wilt vascular staining on avocado (photo by A. Eskalen.)
Redbay Ambrosia Beetle and Laurel Wilt Complex
An invasive ambrosia beetle, the redbay ambrosia beetle (RAB; Xyleborus glabratus) is a serious pest currently spreading through the Florida avocado industry and has been responsible for significant yield reductions since its discovery in 2005. The beetle was first detected in the U.S. in Port Wentworth, Ga. in 2002 and was probably introduced via infested wooden packaging material (Crane 2011).
This beetle has been slowly spreading across the southeastern U.S. and is currently found as far west as east Texas. Therefore, California growers need to be aware that this pest-disease complex may spread to California avocados. What makes this complex dangerous is that the vector, RAB, has a symbiotic relationship with the fungal pathogen Raffaelea lauricola that causes Laurel Wilt Disease (LWD). Native to Southeast Asia, RAB has similarities to our current ambrosia pest, PSHB. However, the fungus associated with LWD is unlike the disease here in California. Trees become infected when female beetles attack host trees and introduce the pathogen into the xylem while boring their galleries. The infection restricts the flow of water in the tree, induces a black discoloration in the outer sapwood and causes the leaves to wilt. Tree mortality is so rapid that leaves do not fall from dying branches.
Symptoms of RAB beetle and LWD infestations include (Carrillo et al. 2017):
Wilting of leaves and young stems. You can usually see dead leaves hanging on branches.
Color change in leaves from light green to dark purplish green or greenish brown.
Stem and limb dieback.
Trunk and major limbs that show dried sap which has a white appearance and is a crystalline, powder-like material.
Dark streaks in the sapwood. Normally sapwood should be white to yellowish white in coloration with no dark staining or streaking. To check for this symptom simply remove a section of the bark to check for discoloration, which, if present, may indicate fungal infection.
Small, dark holes in the sapwood and what looks like loose wood dust and frass indicate wood boring beetles are present.
RAB entry holes in avocado (photo by S. Rios.)
LWD affects redbay (Persea borbonia) and other tree species of the Laurel family (Lauraceae), including avocado. R. lauricola is introduced into host trees when adult RAB colonizes a tree. Adult RAB are very small (~1/16-inch-long), dark brown to black in color and spend most of their life within the tree. Larvae are white, legless grubs with an amber colored head capsule and are found within galleries throughout infected trees. Female beetles can produce flightless male offspring without mating, but females may mate with their male offspring or brothers to produce males and females. Females greatly outnumber males in populations. In the Southeast U.S., RAB’s lifecycle from egg to adult appears to take 50 to 60 days, and there appear to be multiple overlapping generations per year (Hanula et al. 2008). Female beetles emerging from galleries may reinfest the same tree or disperse in search of new hosts. Host trees can remain standing for years and may continue to serve as host material for beetles for several months after initial colonization. Flight activity peaks in the late afternoon and early evening.
It is known that ambrosia beetles are notoriously difficult to control with insecticides because they are protected from residues by living inside the tree most of their life versus being outside the tree. RAB can fly short distances, but LW fungus spreads more quickly through the movement of insect-infested plant material, such as firewood.
Additionally, the pathogen also spreads to other ambrosia beetle vectors. This happens when beetles feed on diseased trees and become contaminated with spores of R. lauricola. Spread can also occur through root grafting between trees. Sanitation is the most effective way to manage this problem. Scouting for wilted branches and quickly removing them has been key to successful early intervention and eradication.
It has been suggested to remove symptomatic trees immediately upon their identification. However, once the appearance of frass and streaks start to show in the wood, this is a sign that the tree has already been infected and has been for some time. As soon as a grower sees the wilt in the branches, it’s time to move quickly. Verticillium wilt and Phytophthora root rot can be mistaken for LW, so avocado growers should check for these diseases before removing trees.
The Florida avocado industry has implemented a universal detection and suppression program with the goal of preventing or limiting the incidence of LW in commercial groves and depressing ambrosia beetle populations (Carrillo et. al 2017). Surveying for the symptoms of LW is a key component to limiting the spread of the disease. Growers and their workers should survey groves weekly or more often if an infestation is detected in an adjacent grove. Early detection, removal and destruction of LW affected trees is the most important practice for controlling LW.
Contact insecticides are ineffective because the pathogen vectors (i.e., the ambrosia beetles) are primarily inside the tree. The first goal is to avoid infestation with the beetles by maintaining a healthy tree as stressed trees are more attractive to colonizing beetles. Beetles prefer trees in orchards that have dense canopies with overlapping leaves and branches.
Chipping infected trees is effective in reducing the spread of the disease. However, chips must be as small as possible (one square inch or smaller) and dried quickly so that the wood is not conducive to fungal growth. A potential drawback to this method is that the aroma of the chipped wood can attract other wood boring insects. Research in Florida is currently being conducted to determine if different commercial formulations of insecticides can be effective in controlling the beetle. For more information regarding LW and RAB, please visit sfyl.ifas.ufl.edu/miami-dade/agriculture/laurel-wilt—a-disease-impacting-avocados/.
Laurel wilt branch die back on avocado (photo by S. Rios.)
Avocado Lacebug
The avocado lacebug (ALB), Pseudacysta perseae (Hemiptera: Tingidae), was first detected in California on backyard avocado trees in Chula Vista and National City, San Diego County in 2004. This pest was first described in Florida in 1908 and has since been reported from Mexico, Guatemala, Puerto Rico, Jamaica, the Dominican Republic and parts of northeastern South America. The native range of ALB is uncertain. Molecular studies suggest that the western areas of Mexico may be the evolutionary center of origin for this pest and it was spread unintentionally from this area, probably on avocado trees into eastern Mexico, Florida and the Caribbean (Rugman-Jones et al. 2012).
After a lull of approximately 13 years, reports started coming of ALB damage to avocados outside of San Diego. In October 2017, well established, reproducing populations of lace bugs were confirmed in commercial Hass avocado groves in Oceanside and De Luz in San Diego County and in Temecula in Riverside County (CAC, 2017). Around this time, infestations were reported from backyard avocados in Culver City in Los Angeles County. Genetic analysis suggested that these new, more damaging populations of ALB were different to those originally detected in San Diego in 2004, and it’s likely that a second introduction of ALB into California has occurred, possibly from Florida (Rugman-Jones and Stouthamer unpublished molecular data).
ALB has been recorded feeding on avocado, red bay and camphor, which are all in the Lauraceae family.
Avocado lace bug is a true bug with sucking mouth parts. Lace bugs use these needle-like mouthparts to feed on the undersides of leaves. Through feeding, leaf cells and pierced cell contents are extracted and ingested, preventing photosynthesis.
Adult avocado lace bugs are small, winged insects, about 2 mm in length (slightly longer than 1/16 in), with black bodies, yellow legs and antennae, and are visible to the naked eye (UC IPM 2021). ALB live in colonies on the lower surfaces of mature leaves, often adults, eggs and nymphs are found together. Eggs are laid in an irregular pattern, sometimes in loose rows, attached to the lower leaf surface and are covered with irregular globules of a black, sticky, tar-like substance excreted by adults. To the naked eye, eggs will appear like grains of black pepper or dirt.
Eggs hatch into wingless nymphs that are capable of walking. Nymphs go through a gradual metamorphosis, shedding their exoskeleton several times as they grow in size, finally developing wings and becoming flying adults. Nymphs are dark red-brown to black and covered with spines. They feed for approximately two to three weeks before maturing into adult males and females which mate, and females then lay eggs, starting the cycle over. In California, ALB populations tend to peak over early summer, around June, then decline in fall, sometime around September (Humeres et al. 2009).
ALB restrict their feeding to the undersides of leaves. Feeding initially causes small white or yellow spots on the surface of the leaves as individual cells dry out. These necrotic areas look like tip-burn caused by excessive salts, but in this case, the necrotic areas are islands of dead tissue in the interior of the leaf surrounded by living tissue. It is suspected that feeding damage can provide entrance for pathogenic fungi, in particular Colletotrichum spp., which are leaf anthracnose fungi. As lace bug colonies grow, brown necrotic areas develop where there has been heavy feeding damage. Heavy feeding can cause striking leaf discoloration and early leaf drop (Hoddle 2004; Hoddle et al. 2005). Avocado lace bug nymphs and adults do not feed on fruit. However, heavy feeding damage to leaves will likely have a detrimental effect on yield, which may result from the loss of photosynthetic capacity in damaged leaves.
Relatively little is known about biology and ecology of ALB in California. In Florida, the most important biological control agents are two egg parasitoids including Oligosita sp. (a Trichogrammatid wasp) and an unidentified mymarid wasp. Green lace wing larvae and other generalist predators are also thought to be important natural enemies. A predatory thrips, Franklinothrips vespiformis, is reported to be the most important natural enemy of the avocado lace bug in the Dominican Republic and is similarly abundant on ALB-infested avocados in Escuintla Guatemala.
Insecticide treatments for other sucking pests currently registered for use on avocado in California will likely provide control of avocado lace bugs (Hoddle et al. 2005). In a trial reported in 1998, J. E. Peña, University of Florida (UF), showed that citrus oil, M-Pede (insecticidal soap) and Mycotrol (a Beauveria fungal species) all controlled lace bug, but it was not indicated how long the effect lasted. Researchers at UF have shown that citrus oil and M-Pede provided short-term lace bug control. Results of small tree trials or weathered residue tests have indicated that carbaryl, imidicloprid, cyfluthrin, carbaryl, fenpropathrin and malathion provide excellent control of avocado lace bug nymphs. Spinosad, abamectin and mineral oil are much less effective at providing control (Humeres et al. 2009b; Byrne et al. 2010). For more information on lace bugs, please visit biocontrol.ucr.edu/avocado-lace-bug.
The presence of frass indicates the presence of RAB (photo by A. Eskalen.)
Proactive Management and Early Detection
The concept of early detection and rapid response is fundamental to effective invasive species management. Developing collaborative relationships with avocado research colleagues in different countries can provide insight into new potential pest problems that could eventually find their way to California (Hoddle et al. 2009). Developing this idea further can result in the development of proactive biological control and IPM programs in advance of the anticipated arrival of new pest species (Hoddle et al. 2018).
Ideally, finding a new pest species in the early stages of the invasion and quickly containing and treating the new infestation may reduce costly long-term management, especially if eradication is possible. Rapid and decisive containment actions are an important consideration as controlling invasive pests, especially fruit feeders, in California avocado orchards will always be a challenge and expensive should they establish widely.
Avocado lace bug adult and eggs (photo by M. Hoddle.)
References
Byrne, F. J., Almanzor, J., Tellez, I., Eskalen, A., Grosman, D. M., & Morse, J. G. 2020. Evaluation of trunk-injected emamectin benzoate as a potential management strategy for Kuroshio shot hole borer in avocado trees. Crop Protection, 132, 105136.
Byrne, F. J., Humeres, E. C., Urena, A. A., Hoddle, M. S., and Morse, J. G. 2010. Field evaluation of systemic imidacloprid for the management of avocado thrips and avocado lace bug in California avocado groves. Pest management science, 66(10), 1129-1136.
Carrillo, D., J. Crane, R. Ploetz, E. Evans, and J. Wasielewski. 2017. Brief Update to Laurel Wilt Recommendations – Ambrosia Beetles. Univ. of Florida, IFAS Extension Notice.
Crane, J. (2011) Laurel Wilt and the Redbay Ambrosia Beetle threaten Florida’s avocado and native trees in the Laurel Family. Univ. of Florida, IFAS Extension Homeowner informational brochure.
Coleman, T. W., Poloni, A. L., Chen, Y., Thu, P. Q., Li, Q., Sun, J., and Seybold, S. J. 2019. Hardwood injury and mortality associated with two shot hole borers, Euwallacea spp., in the invaded region of southern California, USA, and the native region of Southeast Asia. Annals of Forest Science, 76(3), 1-18.
Dowell, R. V., R. J. Gill, D. R. Jeske, and M. S. Hoddle. 2016. Exotic macro-invertebrate invaders in California from 1700 to 2015: An analysis of records. Proceedings of the California Academy of Sciences Series 4 63(3): 63-157.
Hanula, J.L, Mayfield, A.E. III, Fraedrich, S.W., and Rabaglia, R.J. 2008. Biology and host associations of the redbay ambrosia beetle, (Coleoptera: Curculionidae: Scolytinae), exotic vector of laurel wilt killing redbay trees in the southeastern United States. Journal of Economic Entomology 101: pp. 1276-1286.
Hoddle, M. S. 2004. Invasions of leaf feeding arthropods: why are so many new pests attacking California-grown avocados? California Avocado Society Yearbook, 87, 65-81.
Hoddle, Mark S., J. G. Morse, Richard Stouthamer, Eduardo Humeres, Gilsang Jeong, William Roltsch, Gary S. Bender. 2005. “Avocado lace bug in California.” California Avocado Society Yearbook 88: 67-79.
Hoddle, M. S., Arpaia, M. L., and Hofshi, R. 2009. Mitigating invasion threats to the California avocado industry through collaboration. Calif. Avo. Soc. Yrbk., 92, 43-64.
Hoddle, M. S., K. Mace, and J. Steggall. 2018. Proactive biocontrol: A cost effective management option for invasive pests. California Agriculture 72(3): 1-3.
Hughes, M.A., Smith, J.A., Ploetz, R.C., Kendra, P.E., Mayfield, A.B., Hanula, J., Hulcr, J., Stelinski, L.L., Cameron, S., Riggins, J.J. and Carrillo, D. 2015. Recovery plan for laurel wilt on redbay and other forest species caused by Raffaelea lauricola and disseminated by Xyleborus glabratus. Plant Health Progress, 16(4), pp.174-210.
Humeres, E. C., Morse, J. G., Stouthamer, R., Roltsch, W., & Hoddle, M. S. 2009a. Detection surveys and population monitoring for Pseudacysta perseae on avocados in southern California. Florida Entomologist 92(2): 382-385.
Humeres, E. C., Morse, J. G., Stouthamer, R., Roltsch, W., & Hoddle, M. S. 2009b. Evaluation of natural enemies and insecticides for control of Pseudacysta perseae (Hemiptera: Tingidae) on avocados in Southern California. Florida Entomologist, 92(1), 35-42.
Lu, M., Hulcr, J., and Sun, J. 2016. The role of symbiotic microbes in insect invasions. Annual Review of Ecology, Evolution, and Systematics, 47, 487-505.
Mayfield, A.E. III, Barnard, E.L., Smith, J.A., Bernick, S.C. , Eickwort, J.M., and T.J. Dreaden. 2008. Effect of propiconazole on laurel wilt disease development in redbay trees and on the pathogen in vitro. Arboriculture & Urban Forestry 35: 317-324
Mayorquin, J. S., Carrillo, J. D., Twizeyimana, M., Peacock, B. B., Sugino, K. Y., Na, F., and Eskalen, A. 2018. Chemical management of invasive shot hole borer and Fusarium dieback in California sycamore (Platanus racemosa) in southern California. Plant disease, 102(7), 1307-1315.
Mead, F. W., and Peña, J. E. 2016. Avocado lace bug, Pseudacysta perseae (Heidemann)(Insecta: Hemiptera: Tingidae). Entomol. Circ, (346).
Metcalf, R. L. 1995. Invasion of California by exotic insects. California Agriculture 49(1): 2.
Pimentel D, Zuniga R, and Morrison D. 2005.Update on the environmental and economic costs associated with alien-invasive species in the United States. Ecol Econ 52(3):273–288.
Ploetz, R. C., Pérez‐Martínez, J. M., Smith, J. A., Hughes, M., Dreaden, T. J., Inch, S. A., and Fu, Y. 2012. Responses of avocado to laurel wilt, caused by Raffaelea lauricola. Plant Pathology, 61(4), 801-808.
Rios, S., B. Faber, P. Mauk, A. Eskalen, and M.L. Arpaia. 2018. Redbay Ambrosia Beetle Poses Potential Threat to California’s Avocado Industry. California Association of Pest Control Advisors. February 2018. 22(1):36-38, Pp 36-38.
Rivera, M. J., Martini, X., Conover, D., Mafra-Neto, A., Carrillo, D., & Stelinski, L. L. 2020. Evaluation of semiochemical based push-pull strategy for population suppression of ambrosia beetle vectors of laurel wilt disease in avocado. Scientific reports, 10(1), 1-12.
Rugman-Jones, P. F., M. S. Hoddle, P. A. Phillips, G. Jeong, & R. Stouthamer. 2012. Strong genetic structure among populations of the invasive avocado pest Pseudacysta perseae (Heidemann) (Hemiptera: Tingidae) reveals the source of introduced populations. Biological Invasions 14: 1079-1100.
Umeda, C., Eskalen, A., and Paine, T. D. 2016. Polyphagous shot hole borer and Fusarium dieback in California. In Insects and diseases of Mediterranean forest systems (pp. 757-767). Springer, Cham.
Figure 1. Holistic soil management can be used to help improve soil structure and soil quality. We explored the physical and chemical controls in Part 1 and explore the biological controls in Part 2 of this article series (all figures and tables courtesy K. Wyant.)
Management practices that improve soil health and soil quality have gained considerable attention over the past few years. If you are wondering how to get started and what to focus on, you have come to the right place! In Part 2 of this article series, I focus on how the living, biologic components of the soil, the microbes, directly impact your soil, including the structure (e.g., aggregation and pore space). My focus on bacteria and fungi in the soil is a perfect complement to Part 1 of this article series, where we explored the physical and chemical components behind soil structure. Part 1 can be found in the January/February 2021 edition of Progressive Crop Consultant.
Let us kick things off with a quick reminder of some terminology and drive home the connection between soil quality (structure) and soil health (biology).
Soil quality: This term has broad application on your farm. Soil quality refers to how well a soil functions physically, chemically and biologically and does its “job” (Figure 1). Many factors influence the soil quality on a farm and are summed up in Figure 2. In this article, we will focus on the biological management practices that maximize soil quality, expressed here as soil structure.
Figure 2. Crop productivity is influenced by several interrelated concepts, which have an impact on the soil quality of a field.
Soil health: This term refers to the interaction between organisms and their environment in a soil ecosystem and the properties provided by such interactions. When you think of soil health, think of the biological integrity of your field (e.g., microbial population and diversity) and how the soil biology supports plant growth.
There is a direct link between soil health (the living component) and soil quality (the structural component). The linkage is fungi and bacteria in the soil and the byproducts they secrete. These byproducts help restore your soil structure (Figure 3). Furthermore, well-structured soils are characterized by excellent soil health, which indicates a feedback loop between the soil biology and soil particles. But how exactly do the microbes put your soil back together?
Figure 3. The soil on the left has poor soil structure while the soil on the right has excellent aggregation and structure. The samples also have vastly different living components in the soil, as shown in the agar plates. The soil with the best aggregation is characterized by having a healthy soil microbiome (right).
Glue and Nets
Abundant and diverse soil microbial communities produce lots of “free”services for your farm soil. When your soil is healthy, microbes speed up nutrient release rates back to the crop, influence the water holding capacity of the soil, and help restore soil structure. How exactly do they go about sticking the soil together? The answer can be summed up in two terms: glues and nets.
When you have a healthy and abundant soil bacteria population, they produce a sticky, glue-like gel called extracellular polymeric substances (EPS) that forms a protective slime layer around bacteria as they grow. EPS, since it is sticky, acts as a glue for soil particles, sticking them together and improving overall soil structure.
Another important group of soil microbes, called fungi, is most known by its aboveground structures like mushrooms. However, most soil fungi exist where you cannot see them below ground. In the soil, fungi produce millions of miles of microscopic threads called hyphae. These structures help fungi find resources and grow. The threads also help to capture and tie soil particles together like a net, which improves soil structure. Now that you know the connection between your soil biology and soil structure, let us turn our attention to management practices that can help optimize the contribution of microbes to improving your farm soil (Table 1).
Soil Biology Management Guidelines
Feed the Soil Biology
This might seem like an obvious management choice, but this practice is often missed in the yearly crop plan. Your soil is teeming with fungi and bacteria and they are ready to go to work for you. The problem? They are starving and will go dormant on you until conditions improve. Research shows that farm soils are generally low in the food stuffs that microbes like to eat, and that food scarcity will limit the activity of your soil biology. The answer? Provide them with a regular installment of something they like to eat to keep their populations up. You have many choices, including microalgae, molasses, fish emulsions, etc.
Add Soil Biology
Another option is to add biology to the soil. I do not have space to cover all the products out there, but the main concept is to provide selected species of bacteria or fungi to the soil and put them to work for you. Keep in mind that the inoculant must stay alive to get the benefit you are looking for. This can be quite a challenge considering how sensitive microbes are to changes in temperature and humidity from shipping to storage in the farm shop to field application. If you choose this option, make sure your product is viable and high quality when going out into the field.
Mulches and Compost
This practice is like the first management suggestion but provides more of a “slow release” food source for the microbes. Not all the carbon in plant mulch and compost is available as microbial food. Instead, it must be chemically and physically broken down before the microbes can take advantage of it. Another advantage is that mulches and compost provide a nutrient source to the soil. A potential disadvantage here is that mulches and compost can contain excess salts and weed seeds if not prepared correctly.
Reduce Tillage and Improve Soil Biology
Field activities like tillage can be hard on your soil biology, particularly the soil fungi. For example, when a disk moves through the field, it not only slices through the soil (what you want to happen) but it also slices and dices through all the fungi threads you are trying to grow (what you do not want to happen.) This unintentional result can have a severe impact on the biological contribution to soil structure. Moreover, excessive tillage can crush and compact your soil structure, which can set you back from a physical management perspective. Reducing tillage, therefore, can improve your soil structure on two fronts. Talk about a 2-for-1 deal!
Cover Crops and Soil Biology
The cover crop, usually grown in between the rows of permanent crops (e.g., trees and vines) or in the ‘off-season’ for annual crops, can be used to help feed soil microbes. Cover crop roots secrete carbon substances, called exudates, which can help boost the soil fungi and bacteria when a crop is not in the ground. Keeping your soil alive year-round is key to optimizing the biological contribution to soil quality. Fine root hairs can also tie soil particles together, improving soil structure and quality. Another two-for-one deal!
Testing for Soil Biology
No doubt you are familiar with soil tests from your favorite agricultural laboratory. Traditional soil tests have mainly focused on measuring the chemical constituents of the soil (e.g., nitrate, phosphate, etc.) or the physical aspects of the soil (e.g., soil texture, cation exchange capacity). However, as useful as these tests are, they fail to explain how “alive” the soils are. There is good news though! Many laboratories are starting to offer soil health testing services which can help you get a better understanding of the biological components of your soil. Certified Crop Advisors (CCA) help guide growers through which test to order and, more importantly, help them interpret it. Common tests include measurements of carbon dioxide respiration, extraction of DNA for microbial community analysis and even direct counting of fungi and bacteria populations. There are many choices and sound advice from an experienced crop advisor that can help direct you down the right path and reduce the learning curve.
Conclusion
Biological factors can have a profound impact on overall soil structure and, thus, the soil quality of the field. Generally, poorly structured soils have a difficult time supporting optimized crop growth due to the severe reduction in water storage capacity, low oxygen, surface crusting and seed bed issues, accumulation of salinity, etc. If the soil looks like the example on the left side of Figure 3, it may be well worth your time and money to start implementing soil biology improvement practices as outlined in this article and revisit some of the physical and chemical practices discussed in Part 1 of this series. I strongly recommend that you put your field detective hat on to diagnose why your field is not performing as expected. A bit of detective work beforehand can pay off in turning your field around and using your input dollars most effectively.
Dr. Karl Wyant currently serves as the Vice President of Ag Science at Heliae® Agriculture where he oversees the internal and external PhycoTerra® trials, assists with building regenerative agriculture implementation and oversees agronomy training.
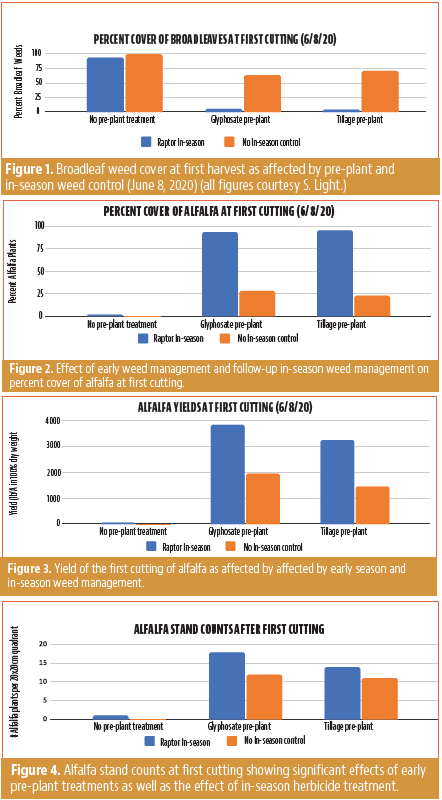
A combination of early weed control combined with in-season weed control was the most successful at controlling weeds and enhancing alfalfa yields.
Good stand establishment is critical for productivity of an alfalfa field both in year one and in subsequent years. Weed competition during stand establishment may be irreversible because it can reduce alfalfa root growth and lead to thinner alfalfa stands and lower forage quality. Thus, it is important to have good weed control during alfalfa stand establishment.
This project evaluated the efficacy of weed control options for both conventional and organic growers. Pre-plant mechanical cultivation and glyphosate spray were evaluated with the goal of providing regionally relevant information about an integrated weed management tool for improved stand establishment.
Methods
Six treatments (Table 1) were replicated three times in the field. Main plots were a pre-plant treatment (either no pre-plant treatment, pre-plant tillage or pre-plant glyphosate). Additionally, half of the plots received later in-season treatment (Table 1): either no treatment or Raptor application in-season after the crop had emerged.
This field was planted in the spring in the Sacramento Valley of California. Weeds were germinated with winter rains. On some plots (treatments 3 and 6), pre-plant glyphosate was sprayed on plots on January 31, 2020 at a rate of three pints glyphosate/acre. On other plots (treatments 2 and 5), mechanical cultivation was implemented on February 11, 2020 once the soil was dry enough. This cultivation was very shallow (top few inches of the soil) to avoid bringing new weed seeds to the soil surface.
Alfalfa seed was flown on the field on March 4, 2020, and the field was then ring-rolled to cover seed and get good seed-to-soil contact. The field was then irrigated for germination a week later. In-season weeds were controlled on some of the plots (treatments 4, 5 and 6) with a tank mix of Raptor (Imazamox Ammonium Salt) at 6 fl oz per acre and Buctril (Bromoxnil) on April 25, 2020.
Data Collected
Baseline weed counts were taken on January 29, 2020 from all plots before treatment implementation but after weed germination. Individual broadleaf weeds and grasses + sedges were counted in three random 20×20 cm quadrats per plot. Plants were counted on this date because weeds and alfalfa plants were small and percent cover would not have captured potential differences.
Weed counts were taken an additional three times between planting and first cutting from all plots. In-season weed counts were taken as percent cover in which the area of the quadrat was broken up in percent covered with broadleaves, grasses + sedges, bare soil and alfalfa. On April 9 and May 14, 2020, weed counts were taken in three random 20×20 cm quadrats per plot, and on June 8, 2020, percent cover was observed in three random square-meter quadrats per plot. The larger quadrat was used for percent cover on June 8 because alfalfa and weeds were tall at this time and the meter by meter square allowed for more accurate representation of each plot.
Plots were hand harvested on June 8, 2020 prior to first cutting by the grower, which occurred on June 10. Two square-meter areas of each plot, which were representative of the larger plot, were cut. Yield biomass was separated into weeds and alfalfa, dried, weighed separately and then converted to a pounds dry matter/acre basis.
Finally, on June 23, 2020, following first cutting, alfalfa stand counts were taken in all plots by counting the number of alfalfa plants in three 20×20 cm quadrats.
Baseline and Early Weed Counts
The first weed counts (January 29, 2020) collected before treatment implementation showed the average count for grasses + sedges for all plots was zero at this count. For broadleaves, there were no significant differences by treatment, but there were significantly more weeds in the side of the field with no in-season control compared to the side where Raptor was applied in-season.
April 9, 2020 Weed Counts
Grasses + sedges: There were not many grasses or sedges in the field.
Broadleaves: There were significantly less broadleaves in the plots that had pre-plant weed control (glyphosate or tillage).
Alfalfa: Alfalfa plants were small at this counting date; however, there were significant treatment differences with the pre-plant weed control treatments having more alfalfa than the control.
May 14, 2020 Weed Counts*
Grasses + sedges: There were not many grasses or sedges in the field.
Broadleaves: There were significantly less broadleaves in the plots that had pre-plant weed control (glyphosate or tillage) and in the plots that had Raptor applied in-season.
Alfalfa: There was significantly more alfalfa in the plots that had pre-plant weed control (glyphosate or tillage) and in the plots that had an in-season herbicide.
*Data not shown
This photo taken at harvest shows how heavy the weed pressure was even in plots with glyphosate or tillage pre-plant that did not have in-season herbicide application.
Broadleaf Weeds Dominated at First Cutting
There were significantly more broadleaf weeds in the plots that had no pre-plant weed control (glyphosate or tillage) (Figure 1). Additionally, the plots that had Raptor applied in-season reduced broadleaf weeds down to negligible levels compared with no in-season treatment (Figure 1). There were not many grasses or sedges in the field; however, there were more grasses in the side of the field with no in-season herbicide application.
Alfalfa Stand
There was significantly more alfalfa at first cutting in the plots that had pre-plant weed control (glyphosate or tillage) and in the plots that had an in-season herbicide (Figure 2). Weeds in the no pre-plant treatment essentially killed many of the young seedlings due to weed competition. This is a key issue since early growth and establishment of alfalfa seedlings sets the stage for vigorous growth over many years of production. This is demonstrated by the number of alfalfa plants in a 20×20 cm quadrant after first cutting (Figure 4). There were significant differences in the alfalfa stand after first cutting. With regard to pre-plant treatments, both glyphosate spray and tillage pre-plant significantly increased alfalfa stand compared to the plots with no pre-plant treatment.
Enhanced Yields
Alfalfa yields were near zero for the plots where early control was not applied (Figure 4). Additionally, yields were improved over 90% when an in-season weed control was applied (Figure 4). This yield data is only for the first cutting of the stand, not for the full first year of production. There were significant differences in alfalfa yield between pre-plant treatments and plots that had no pre-plant weed control (Figure 3). Both the glyphosate and tillage pre-plant treatments increased yields. In addition, the Raptor spray significantly increased yields compared to plots without in-season control.
A combination of early weed control combined with in-season weed control was the most successful at controlling weeds and enhancing alfalfa yields.
Biomass was separated into alfalfa (Figure 3) and weeds after plots were hand-harvested. Alfalfa and weeds were then weighed separately by plot. There were significantly more weeds by weight in the side of the field that did not get the herbicide spray in season compared to the side that did get an herbicide spray. However, within one side of the field (Raptor or not), there were no significant differences by pre-plant treatment. In other words, even though there was more alfalfa in the plots with pre-plant weed control, there were also more weeds. The photo taken at harvest show how heavy the weed pressure was even in plots with Glyphosate and tillage pre-plant that did not have in-season herbicide application.
When comparing plots with the same pre-plant treatments with or without in-season herbicide spray, plots that were tilled pre-plant did not have significantly different stand counts regardless of in-season herbicide treatment. However, within the plots that were sprayed with glyphosate pre-plant, those that also were sprayed with Raptor in-season had significantly higher alfalfa stand counts than those without in-season control.
Conclusions
The data shows that controlling weeds prior to planting, either with shallow tillage or an herbicide spray (glyphosate), will reduce weed pressure, increase yields and lead to a stronger alfalfa stand after first cutting. There were also differences between plots that got an in-season herbicide and those that did not. Yields were highest in plots that had both pre-plant weed control and an in-season herbicide. The plots with the highest stand counts after first cutting were also the plots that had both pre-plant and in-season weed control. However, the stand in the pre-plant treatment plots that did not have an in-season herbicide application still had relatively high alfalfa stand counts after first cutting. This means that with early effective weed control, the alfalfa stand may be more robust for future cuttings, even if weed pressure was high initially. As shown in the provided photos, the alfalfa was robust in the understory of the canopy, even when broadleaf weeds were very large. By first cutting, many broad leaf weeds had gone to flower, so they likely would not return after first cutting. However, when included in the harvest, these weeds reduce quality and price of the hay and also contribute seed to the weed-seed population in the field.
Ideally, both pre-plant and in-season weed control would be implemented to get highest yields, quality, a vigorous stand and ensure animal safety. However, growers (particularly organic) may be able to do a pre-plant tillage to control weeds and establish a good alfalfa stand, accept some yield reduction and additional weed pressure leading up to first cutting and then have a strong alfalfa stand for subsequent cuttings.
The author would like to thank the California Alfalfa & Forage Association for funding this project and River Garden Farms for their collaboration on this project
Stand count data was collected after first cutting (see results in Figure 4.)










































 Invading California")

