









Nitrogen (N) is one of the most managed nutrients in vineyards, since it strongly affects vine growth and fruit development. Although numerous studies have been conducted to understand how N fertilization influences vine productivity and fruit composition, the impacts of N on vine nutrient status and soil microbes receive less attention.
Among a wide range of soil microbes that play vital roles in soil health and vine productivity, arbuscular mycorrhizal fungi (AMF) are unique due to their symbiotic association with grapevines and their contribution to vine nutrient acquisition. Arbuscular mycorrhizal fungi obtain nutrients from the soil, especially for phosphorus (P) and other poorly mobile nutrients, and transfer those nutrients to the plant. In turn, plants provide sucrose and fatty acids to AMF to support fungal functions and growth.
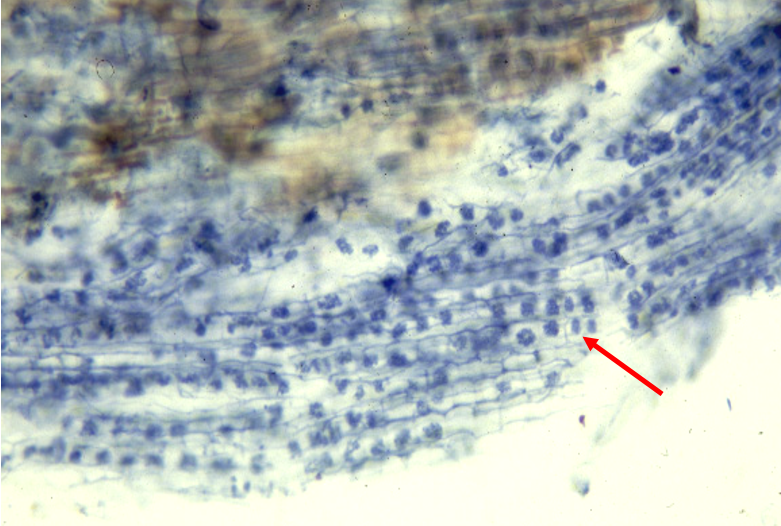
To sustain intensive nutrient exchange between two partners, AMF colonize individual cortical cells of fine roots and form arbuscules, which are tree-like fungal structures that greatly increase surface area contact between plants and AMF (Fig. 1). Grapevines are considered a “super” host of AMF. The percentage of fine roots colonized by AMF is generally above 60% in field and greenhouse conditions. Such high colonization rates also reflect the great dependency of grapevines on AMF. Indeed, non-mycorrhizal vines are stunted in low P soils, while mycorrhizal vines could acquire adequate P from the soil, overcome P limitation and grow normally.
Willamette Valley Trials
Maintaining AMF colonization in grapevine roots is particularly important for vineyards in Oregon’s Willamette Valley where red-hill soils are most commonly found. Since those highly weathered acid soils have low P availability, grapevines rely on AMF for ample P acquisition. In other crops, N fertilization was shown to reduce root colonization by AMF, but it is unclear whether N applied at moderate rates would decrease mycorrhizal colonization in grapevines. If N applications would suppress AMF and impair vine P uptake, this negative effect should be accounted for when developing fertilization management plans for vineyards. This article summarizes part of my Ph.D. research conducted in Oregon, which explored how vineyard N applications affect vine nutrient status, root growth and AMF.
Experiments were conducted in a Chardonnay vineyard and a Pinot noir vineyard over three years in Willamette Valley. Both vineyards are somewhat limited by N but have varying levels of soil P. At each site, we evaluated three treatments, including no N application (No N), N applied to the soil (soil N) and N applied to the foliage (foliar N). Each treatment was replicated four times. The soil N vines were fertilized two or three times between bud break and veraison using UAN-32 at the rate of 40 to 60 lbs N/acre/year. The foliar N vines received three urea sprays to the canopy from fruit set to two weeks post veraison at the rate of 19 to 23 lbs N/acre/year.
All treatments were evaluated across three years in both vineyards with the exception that foliar N treatment was assessed only in Year 2 and 3 in Chardonnay. In each season, leaf blades and petioles were sampled at bloom and veraison for nutrient analysis. Due to the late initiation of foliar N treatment in Chardonnay, bloom leaf samples were collected only in Year 3 for this specific treatment. Soils and roots were sampled three times a year when berries were pea-size, near veraison and about a month after harvest.
Effects of Soil N Applications in Chardonnay
As expected, soil N applications increased vine N status starting from Year 1 (Fig. 2). For simplicity, only petiole nutrient data at bloom are presented here. Changes of nutrients in corresponding leaf blades followed a similar trend. Previous work on Pinot noir in the Willamette Valley proposed 0.7% as the critical value of petiole N concentration at bloom to ensure sufficient yield (2.5 to 3.5 U.S. ton/acre) and adequate fruit N.
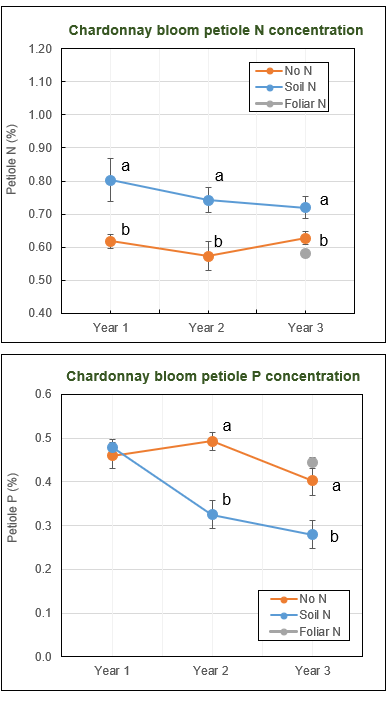
Since Chardonnay vines in this region generally carry heavier crop load (four to five U.S. ton/acre) and develop larger canopies compared to Pinot noir vines, Chardonnay vines may have a higher N requirement. With a petiole N concentration of 0.6% at bloom, Chardonnay vines that received no N applications clearly experienced some N limitation in this study. Soil N applications improved bloom petiole N concentration by 15% to 30%. In response to greater vine N status, the soil N vines had 30% higher yield and about 35% more pruning mass as compared to the no N vines.
Soil N applications decreased vine P status in Chardonnay in Year 2 and 3, where petiole P concentration at bloom was about 30% lower in the soil N vines than no N vines (Fig. 2). The negative effect of soil N fertilization on vine P status became more evident in late season. The concentration of petiole P at veraison decreased 50% in the soil N vines in the last two years of the experiment.
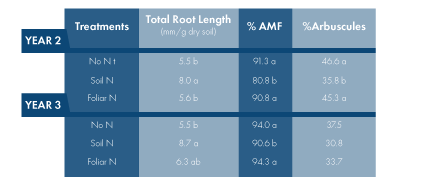
Why did soil N applications reduce vine P status? The most straightforward answer would be the dilution of P in leaves due to N stimulated canopy growth. However, this is unlikely the sole reason. Soil N applications increased veraison leaf area by 10% to 19% in Year 2 and 3, while the corresponding leaf blade P decreased to a larger extent (19% to 29%). The second possible explanation for the decreased leaf P under increased soil N supply is that fertilization altered P allocation within the plant and less P was translated above ground. Indeed, because soil N fertilization increased root growth (Table 1), more P can be retained belowground to support new root development. Yet, this assumption is not supported by our observation in the greenhouse or previous studies where soil N fertilization generally increases the proportion of P allocated to aboveground tissues. Unfortunately, we did not sample roots for nutrient analysis in this study, and thus effects of soil N on root P concentration cannot be further examined. In addition to the two explanations presented above, we suspect that soil N applications might lower AMF colonization in roots and therefore decrease vine P uptake.
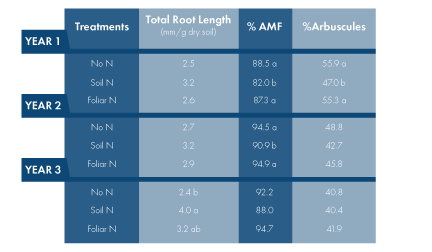
The percentage of fine roots colonized by AMF (fungal hyphae, arbuscules, vesicles and spores) decreased with soil N supply in Year 2 and 3 (Table 1), in accordance with reduced vine P status in Chardonnay. The percentage of roots colonized by arbuscules also reduced in the soil N vines in Year 2, but not in Year 3. The greater suppression of arbuscular colonization in the soil N vines in Year 2 was likely attributed to the fact that more N (20 lbs N/acre) was applied in Year 2 than Year 3. Clearly, increased root growth played a role in the decrease of AMF in the soil N vines because root colonization usually lags behind root growth. Soil N fertilization might affect AMF through other mechanisms as well. For example, N fertilization could reduce the amount of carbon translocated from vines to AMF and, in turn, decrease P delivered by the fungus. Or soil N supply reduced N translocated from AMF to vines, resulting in a decrease of mycorrhizal colonization.
Effect of Soil N Applications in Pinot Noir
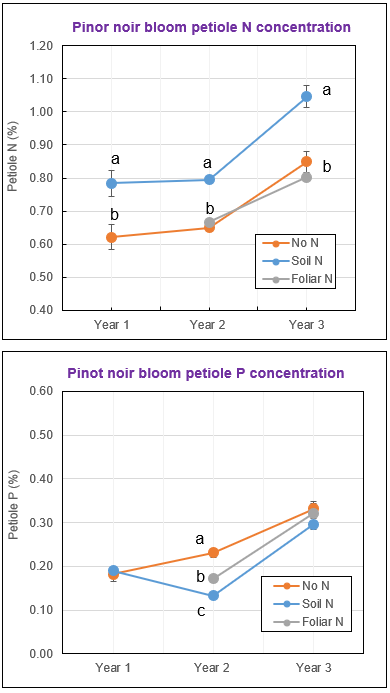
Similar to what we observed in Chardonnay, soil N applications improved vine N status in Pinot noir across three years (Fig. 3). Soil N fertilization also increased root growth and decreased mycorrhizal colonization in Pinot noir, although the effects were less evident as compared to Chardonnay (Table 2). The percentage of roots colonized by AMF was lower in the soil N vines than no N vines in two of three years, while the percentage of roots colonized by arbuscules reduced in the soil N vines only in one year.
Even though soil N altered mycorrhizal colonization, it had no influence on leaf blade or petiole P concentration at bloom or veraison in any year, except petiole P concentration at bloom was lower in the soil N vines than no N vines in Year 2 (Fig. 3). Even so, P concentration of corresponding leaf blades was not affected by soil N supply, suggesting an overall small impact of N fertilization on vine P status in Pinot noir. The difference in how vine nutrition and mycorrhizal colonization responded to soil N application between Chardonnay and Pinot noir can be attributed to the difference in soil N and P availability.
Compared to the Pinot noir vineyard, the Chardonnay vineyard has lower soil N and higher P concentration. It seems soil N fertilization would suppress AMF colonization and decrease vine P status to a greater extent in vineyards with lower N and higher P availability.
However, since the Chardonnay and Pinot noir vineyards differ in many other aspects, such as canopy size, irrigation and cropping level, the comparison between these two varieties are not straightforward. Thus, upon the completion of field experiments, we conducted a series of greenhouse experiments to further examine how N and P regulate vine nutrient status and mycorrhizal colonization under a more controlled environment. The negative effect of soil N applications on AMF was observed again in vines supplied with N at a high rate in the greenhouse.
Effect of Foliar N in Chardonnay and Pinot Noir
Foliar N applications had minor influence on vine N status, vine P status, root growth, and mycorrhizal colonization in both varieties (Figs. 2 and 3, Tables 1 and 2). This is somewhat expected, since a large amount of N applied to the foliage appeared to be transferred to the fruit rather than other plant organs.
Conclusions
The evidence obtained from the field experiments indicates that soil N fertilization at moderate rates can negatively influence mycorrhizal colonization and reduces the benefits conveyed by this symbiotic relationship. Foliar N applications, on the other hand, had no impact on AMF or vine P status. The negative effect of soil N applications on mycorrhizal association provides another justification for being judicious with N fertilization in vineyards.
This project was funded by Oregon Wine Board and USDA-ARS. The author would like to thank Erath winery and Results Partner Inc. for their help and support.






 Invading California")


{kind=link}