

Irrigation
The Agronomy of Water
Water ties all aspects of crop production together and is the largest input on most farm operations on…
Read ArticleArticle Archive
Water ties all aspects of crop production together and is the largest input on most farm operations on…
Read Article
It is estimated that approximately 78 million acres in the western San Joaquin Valley are affected by soil…
Read Article
The goal of properly timed foliar-applied fertilizer is to increase the economic benefit derived from the grower’s fertilization…
Read Article
In California, avocados are primarily grown in southern and central parts of the state along the coast. These…
Read Article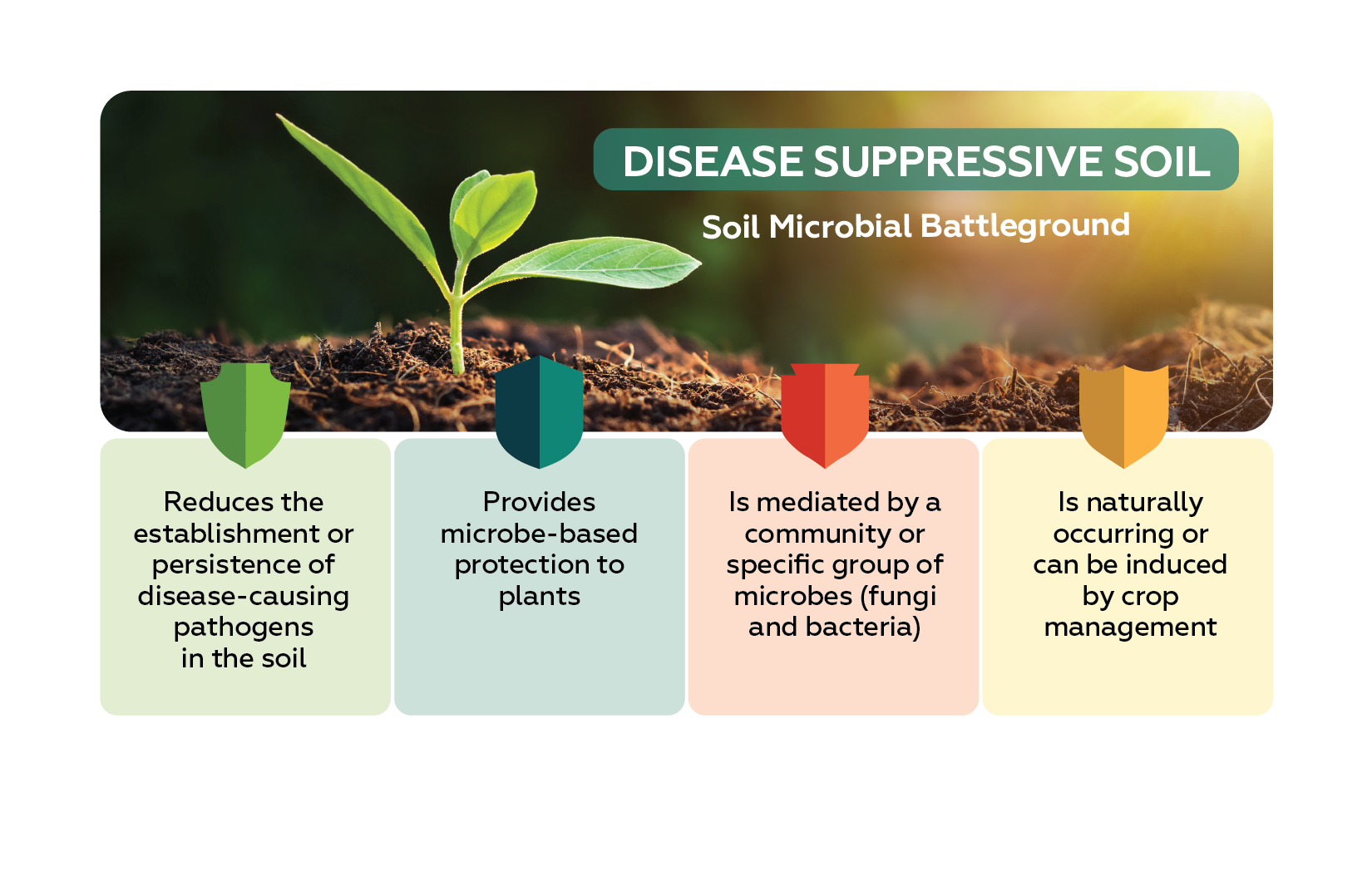
When it comes to managing soilborne plant diseases, methods for reducing or eliminating the impact of pathogens have…
Read Article
Phosphite has been a controversial topic for years. Its use and benefits are argued in hundreds of research…
Read Article
Postharvest scouting for weeds in tree nut orchards presents an opportunity to evaluate this year’s orchard floor management…
Read Article
Citrus tree growth, health and productivity are influenced by the rootstock chosen. In a Citrus Research Board webinar,…
Read Article
Mealybug management is an important part in controlling grapevine leafroll disease. At a Fresno State grapevine viruses symposium,…
Read Article
An unusual strain of beet curly top virus (BCTV) has been confirmed in Colusa County this growing season.…
Read Article