

Agriculture
New Tool Brings Data-Driven Cover Crop Recommendations to the West
There is a new cover crop tool available for growers, technical assistance providers, crop advisors and other users…
Read ArticleArticle Archive
There is a new cover crop tool available for growers, technical assistance providers, crop advisors and other users…
Read Article
One of my favorite activities to do with my kids is going to the movies. They’re at a…
Read Article
When a fertilizer label lists nitrogen, phosphorus or potassium, growers know those numbers mean something. They’re backed by…
Read Article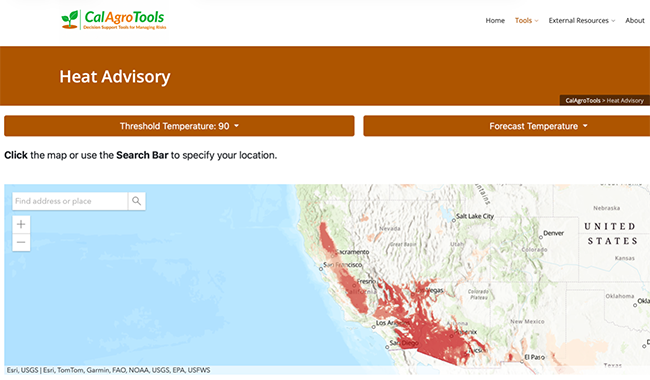
Abstract Weather and climate variability has direct influence on agricultural production systems. While weather data are typically freely…
Read Article
Rising fertilizer prices and increasing regulatory pressure push growers throughout California to consider decreasing mineral N use. Many…
Read Article
Background 4R nutrient stewardship (right rate, time, source and placement) is central to nitrogen (N) management, but how…
Read Article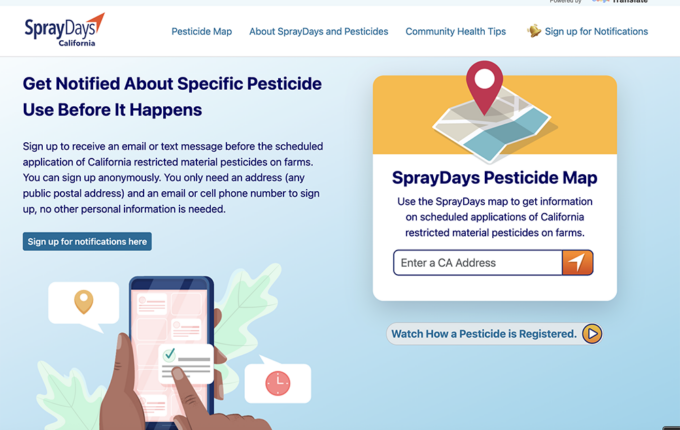
Over the past several months, I have been paying close attention to the growing conversation surrounding paraquat in…
Read Article
Pest Tactic Journal Provides Open-Access Platform for Pest Control Research Pest Tactic Journal was established a few years…
Read Article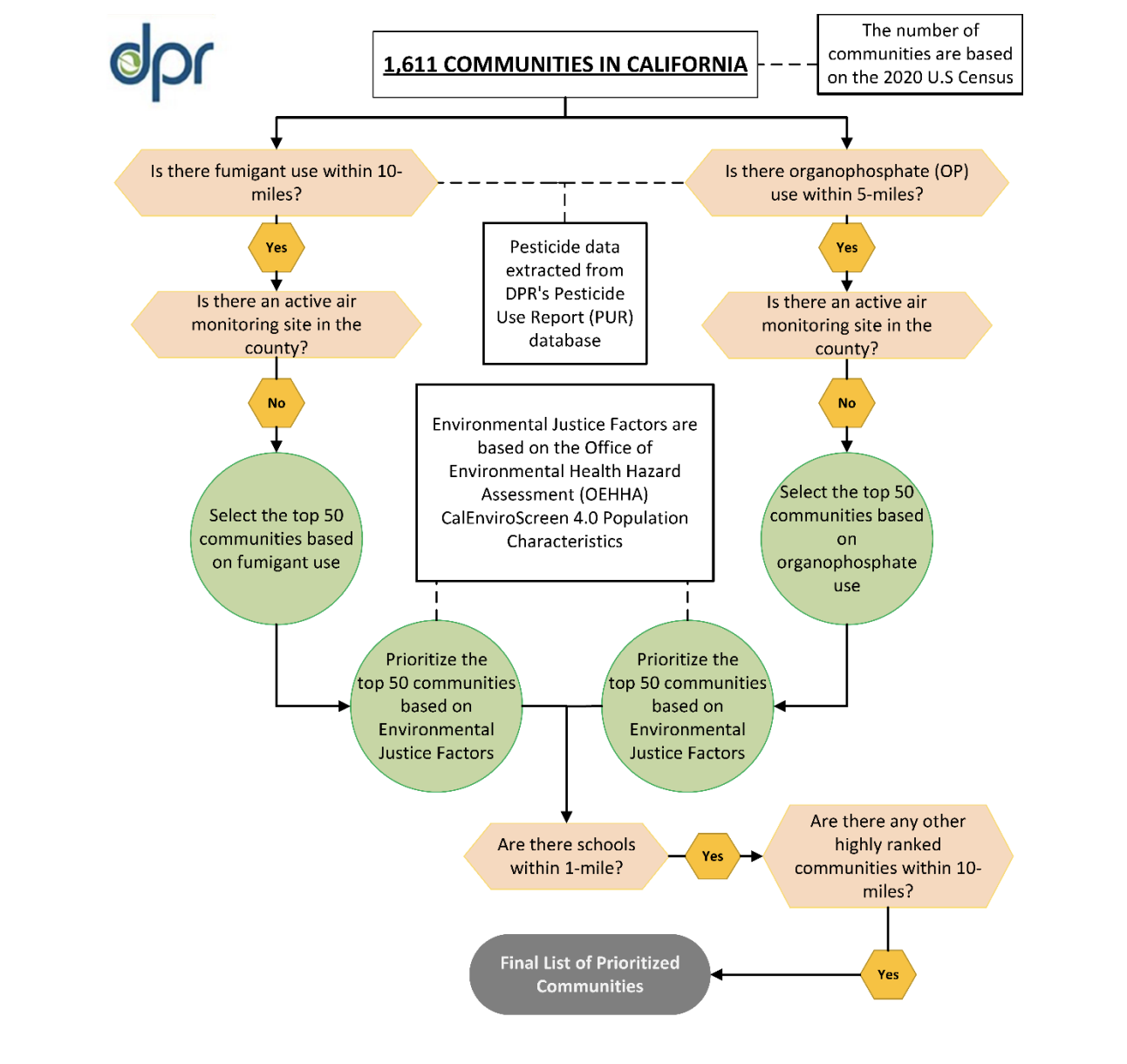
The California Department of Pesticide Regulation (DPR) has announced plans to expand its statewide Air Monitoring Network (AMN).…
Read Article
Lettuce Fusarium Wilt Fusarium wilt has become one of the most challenging diseases for California lettuce growers. Once…
Read Article