

Grapes
Effect of Heat on Grapevine Production and Fruit Quality
The San Joaquin Valley (SJV) is already considered a hot growing region for winegrapes, and heat stress is…
Read ArticleArticle Archive
The San Joaquin Valley (SJV) is already considered a hot growing region for winegrapes, and heat stress is…
Read Article
Developing a weed management program in conventional pear orchards is a challenge, varies from orchard to orchard and…
Read Article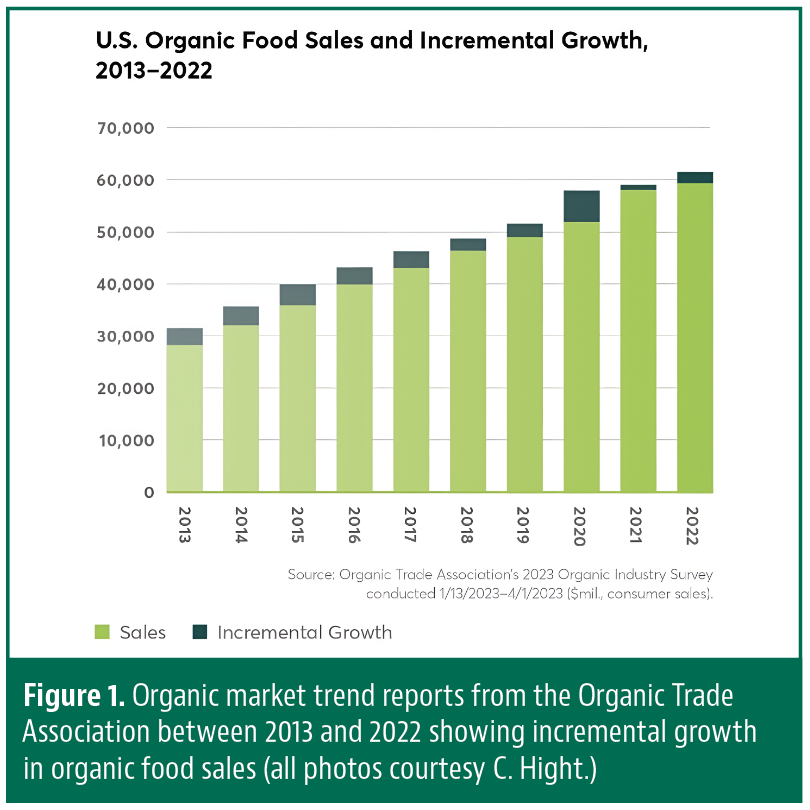
The first concepts of organic agriculture as we now know it were developed in the early 1900s by…
Read Article
Many California specialty crop production areas often face significant water supply curtailments due to recurring droughts and stringent…
Read Article
In the Western U.S., alfalfa weevil is one of the key arthropod pests. Left unmanaged, it can defoliate…
Read Article
Growers will need to implement practices to protect groundwater from nitrate contamination to comply with the water quality…
Read Article
As we kick off 2025, it’s clear agriculture technology is not just a buzzword; it’s the lifeblood of…
Read Article
Over 90% of all almond acres farmed in California are using fertigation to deliver crop nutrition. How-ever, the…
Read Article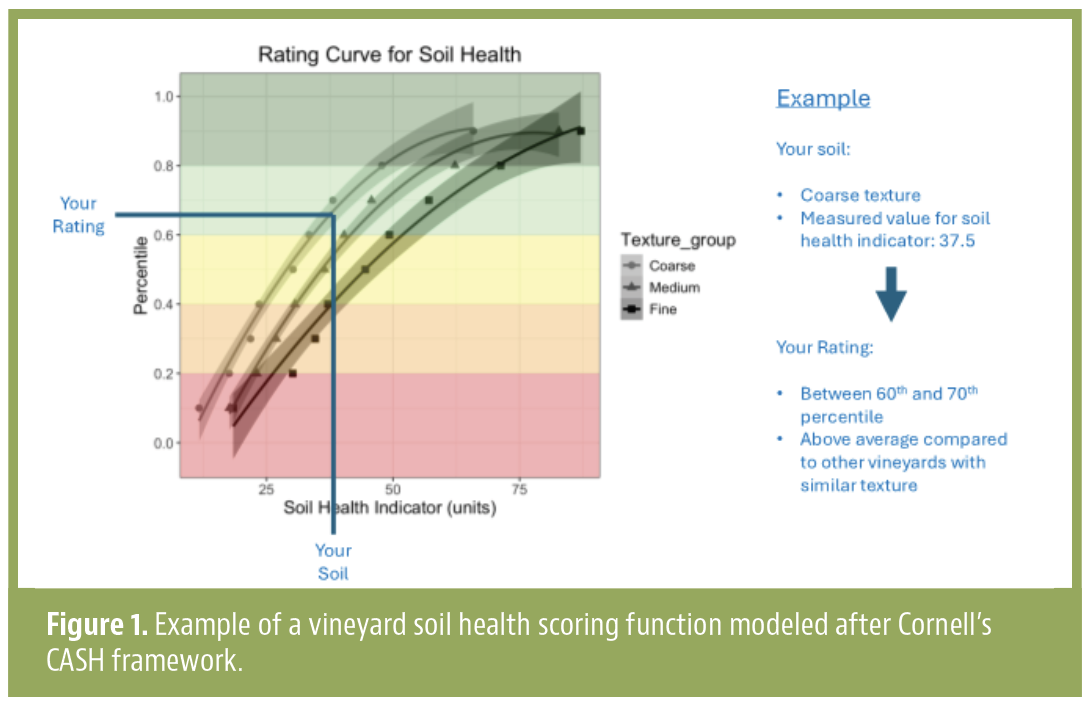
Soil health is central to sustainable agriculture and a key goal of regenerative and organic farming. Practices like…
Read Article
Dry root rot, caused by the soilborne fungus Fusarium solani, has been a persistent threat to California…
Read Article