









As we kick off 2025, it’s clear agriculture technology is not just a buzzword; it’s the lifeblood of modern farming. Every year, we at Progressive Crop Consultant witness an influx of innovations designed to empower consultants, optimize crop production and meet the challenges of a rapidly changing agricultural landscape. This year is no exception.
For those of you in the field walking orchards, inspecting vineyards or evaluating broad-acre crops, you know firsthand staying informed about ag tech advancements isn’t optional. Your clients depend on you for insights that can boost yields, reduce costs and navigate increasing environmental and regulatory pressures. Let’s explore the most impactful ag tech trends shaping the industry in 2025 and how they can elevate your role as a trusted crop consultant.
Precision Agriculture: The Next Generation
Precision agriculture has been a mainstay in ag tech for years, but in 2025, it’s entering a new era. Today’s tools are faster, smarter and more precise thanks to artificial intelligence (AI), machine learning and next-generation IoT devices.
From drone imaging systems that provide real-time, multispectral field data to IoT-enabled sensors monitoring soil health, these technologies are making your job as a consultant more efficient and accurate. AI platforms now analyze complex datasets instantly, recommending specific actions like targeted fertilization, herbicide applications or irrigation adjustments. These are no longer nice-to-have tools; they’re critical for making data-driven decisions that save clients time and money.
What’s exciting is how accessible these tools have become. Many of your clients may already have access to precision ag hardware or platforms like FieldView or AgLeader. As their consultant, your ability to interpret and act on the outputs of these technologies could be the difference between an average season and a bumper crop.
The Soil Health Renaissance
Soil health has always been a cornerstone of successful farming, but 2025 is taking soil science to a new level. Digital soil mapping and biological inputs are the tools of choice this year.
Today’s digital soil maps provide real-time data on compaction, nutrient availability and microbial activity, giving consultants granular information that would have taken weeks to compile a decade ago. This precision allows you to recommend site-specific practices like micro-dosing nutrients, deep tilling, or even introducing cover crops.
On the biological side, engineered microbial solutions and biostimulants are offering new ways to improve nutrient uptake and resilience. These products are more advanced than ever, often tailored to specific crops, soil types or climate conditions. If you’re not already exploring biological inputs with your clients, now is the time; they’re a game-changer for sustainable and profitable farming.
Automating Pest and Disease Management
Managing pests and diseases is often one of the most time-consuming and costly aspects of farming. Thankfully, automation is stepping up in 2025.
Drones equipped with thermal imaging cameras can now identify hotspots for pests or diseases before they become widespread. Meanwhile, autonomous sprayers can target those areas with precision, reducing chemical use and labor costs.
Even better, AI-based pest ID apps allow you to take a picture in the field and get immediate recommendations for treatment. These tools are invaluable for consultants managing diverse crops or large acreage. When combined with historical pest data and weather patterns, you can offer clients a predictive pest control strategy rather than reactive solutions.
Climate Resilience: Meeting the Moment
Let’s face it: Climate change is no longer a theoretical problem. Droughts, heatwaves and erratic weather are reshaping agriculture. As consultants, your ability to guide clients through these challenges is more important than ever.
Ag tech in 2025 offers a host of climate resilience tools. Smart irrigation systems, for example, use weather forecasts and real-time soil moisture data to optimize water usage. Predictive modeling software can simulate how specific crops will perform under different weather scenarios, helping you recommend the right varieties and planting schedules.
Perhaps most exciting are the advances in heat-tolerant and drought-resistant crop genetics. These breakthroughs provide a lifeline for growers in regions where water scarcity or temperature extremes threaten yields. As their consultant, you’re in the perfect position to guide clients on integrating these new technologies into their operations.
Data Integration and Decision-Making
One of the biggest challenges we hear from consultants is “data overload.” With sensors, drones and apps collecting data from every angle, how do you sort through it all?
Thankfully, farm management platforms are becoming more sophisticated. Tools like Agworld and Granular integrate data from multiple sources (e.g., yield monitors, soil sensors, financial records) into a single dashboard. This makes it easier for consultants to identify patterns and offer actionable recommendations.
For those of you working with clients in premium markets, blockchain technology is also worth exploring. Blockchain ensures traceability, helping growers prove their crops meet organic or sustainable standards. As a consultant, helping clients implement traceability systems can open doors to higher-value markets.
Sustainability as a Profit Driver
We’ve talked about sustainability for years, but in 2025, it’s becoming a revenue generator. From carbon markets to regenerative agriculture, sustainability practices are now being financially incentivized.
Carbon credit programs, for example, reward growers for practices like cover cropping or reduced tillage. Consultants play a key role in helping clients implement these systems and navigate the application process.
Similarly, the growing demand for ecofriendly inputs is creating opportunities for consultants to recommend alternatives that align with market trends. Sustainable farming is no longer just good for the planet; it’s good for the bottom line.
If there’s one thing we’ve learned in 2025, it’s ag tech is here to stay. But while the tools are powerful, they’re only as effective as the people who use them. That’s where crop consultants come in.
You’re not just a service provider; you’re a trusted advisor, a problem solver and now a technology integrator. The role you play in interpreting and implementing these tools is more critical than ever. By staying informed and embracing these innovations, you ensure not only your clients’ success but also the future of agriculture itself.
At Progressive Crop Consultant, we’re committed to keeping you informed about the latest trends, tools and techniques. Together, we can tackle the challenges of 2025 and build a more resilient, profitable future for agriculture.
Let’s make this year one for the record books.


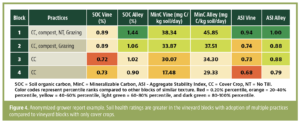

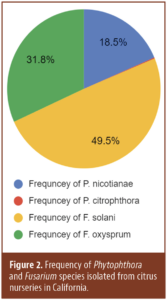
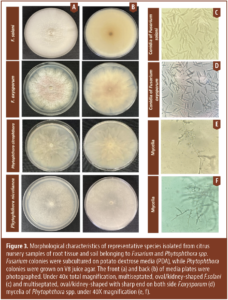

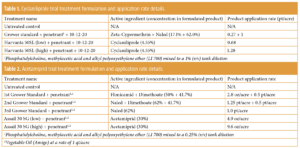
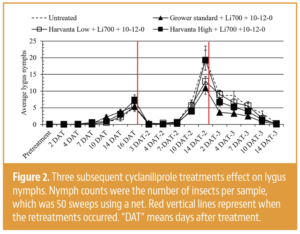
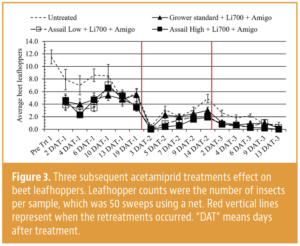




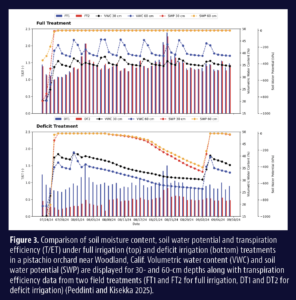


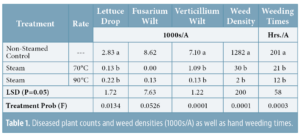











 Invading California")

