









In 1905, German scientist Fritz Haber successfully synthesized ammonia from atmospheric nitrogen gas, an inert form of nitrogen resistant to chemical and biological reactions. Shortly thereafter, BASF chemist Carl Bosch built upon Haber’s work to facilitate large-scale ammonia synthesis, and the process was dubbed Haber-Bosch N fixation. Although synthetic N fixation was initially developed for chemical weapons production, the technology was soon turned toward fertilizer manufacturing.
Suddenly, the essential nutrient most limiting to crop production became available in abundance, and the world’s food supply soared. Haber-Bosch N fixation revolutionized agriculture, but the readily available N fertilizer came at a cost to surface and groundwater resources. Excessive nitrate in farm water runoff disrupts aquatic ecosystems, and nitrate leached down the soil profile to aquifers renders well water unsafe for human and livestock consumption. Increasingly stringent water quality regulations motivate growers to look for new N management practices, and recent research suggests that beneficial N-fixing bacteria could provide a valuable solution.
Synthetic N fixation is a relatively new and disruptive human technology, but microorganisms have relied on biological N fixation for millennia without endangering the surrounding environment. Several types of bacteria, including the well-known rhizobia associated with legumes, convert atmospheric N into plant-available ammonium N. Most ag professionals are familiar with leguminous rhizobia, but there are many other types of bacteria that can contribute N to a wider range of crops.
N-fixing bacteria can be categorized into three main groups: symbiotic, associative and free-living species. Symbiotic N-fixing bacteria live inside plant cells, usually within special organs like root nodules. Associative bacteria live on plant surfaces or in interstitial spaces, while free-living bacteria inhabit rhizosphere soil but do not require direct contact with plant roots. Bacteria in each category fix N for their own use, and plants nearby benefit from the excess. Symbiotic bacteria directly deliver N to the plant, while associative and free-living organisms release bioavailable N at variable rates, depending on soil N concentration and other environmental conditions.
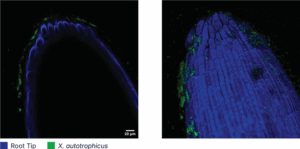
Symbiotic Nitrogen Fixers
Symbiotic N-fixing bacteria form a mutually beneficial relationship with plants by providing ammonium in exchange for a carbohydrate-rich energy source. Symbiotic N-fixing partners include rhizobia with legumes, Frankia with alder, and cyanobacteria with cycads. Rhizobia and their leguminous hosts are well researched and commonly used in agriculture, contributing between 80 to 300 pounds N per acre when effective nodulation is achieved. Individual rhizobia species and strains are adapted to coexist with specific legumes, and some partnerships are more effective than others. For example, Sinorhizobium meliloti provides alfalfa with 200 to 300 pounds N per acre, meeting most if not all the crop’s growth requirement. Conversely, garbanzo beans colonized with Bradyrhizobium japonicum receive just 25 to 80 pounds N per acre through atmospheric fixation. Biological N fixation potential is usually high in legumes but depends on successful nodulation and favorable environmental conditions. Rhizobia depend on their plant host for survival, so conditions that stunt crop growth, such as drought, extreme temperatures or pathogen pressure, will also reduce N fixation. Growers can get the best chance at maximizing N fixation by inoculating seeds with the right rhizobia strain to match their crop. Select a formulation that includes a sticker with a carbohydrate food source to keep the bacteria well fed and close to the seed until roots have grown enough to enable nodulation. Inoculants should be stored at room temperature out of direct sunlight and applied to seed less than 24 hours before planting to maintain viability.
‘…it is clear that under the right circumstances, [nitrogen]-fixing bacteria can help growers produce healthy crops and offset [nitrogen] fertilizer requirements.’
Associative Nitrogen Fixers
Legumes accommodate rhizobia by growing nodules to host the bacterium, but other crops can benefit from biological N fixation without changing their physiology. While symbionts like rhizobia live inside plant cells, associative microbes colonize the surfaces of roots and foliage or live in interstitial spaces between plant cells. Many of these N fixers associate primarily with grasses, such as wheat, corn and rice, but some may be compatible with a wider range of plant types. Researchers have observed that associative bacteria such as Azospirillum, Bacillus and Gluconobacter can provide roughly 20% to 25% of the N requirement in both corn and rice. One study showed that inoculating corn with Pseudomonas and Bacillus megaterium resulted in equal crop dry weight and chlorophyll concentration as applying 33% N fertilizer. Sugarcane colonized by Gluconacetobacter diazotrophicus may receive an even higher N supply at over 150 pounds N per acre, according to a report from North Dakota State University. Research investigating the biological links between associative bacteria and corn found that a particular variety grown in Mexico promotes microbial colonization and N fixation by excreting mucilage on aerial roots to attract associative microbes and create the anaerobic environment required for N fixation. These observations generate interest in breeding crop varieties better adapted to facilitate biological N fixation and reduce reliance on N fertilizer.

Free-Living Nitrogen Fixers
N-fixing bacteria that live in the soil and do not require direct contact with plants can also offset N fertilizer requirements, given the right environmental conditions. N fixation requires anaerobic conditions because the key nitrogenase enzymes involved in the process cannot function in the presence of oxygen. Most free-living bacteria successfully fix N in saturated soils or when they find anaerobic microenvironments in otherwise aerated soil. Several types of bacteria including Clostridium, Herbaspirillum and Azospirillum thrive in flooded soils such as rice paddy fields, fixing enough N for their own use while meeting some of the crop’s N demand as well.
Not all free-living N fixers require a field. Bacteria in the genus Azotobacter live in aerated soils and fix atmospheric N even in an oxygen-rich environment. Azotobacter chroococcum, A. vinelandii and other aerobic species are available as soil inoculants. Studies suggest that they usually fix around 20 pounds N per acre per year, but one study reported a much higher N fixation rate of over 50 pounds per acre per year. Sometimes these bacteria fix more than enough N for their own use, allowing plants to share the excess. More N is released as bacterial populations die and turn over, allowing other soil microorganisms to mineralize the N stored in their bodies into plant-available forms.
Free-living soil bacteria may not provide as much plant-available N as symbiotic and associative bacteria, but they benefit plants in other ways. Azotobacter and Azospirillum species are well known for stimulating crop growth by secreting plant hormones such as cytokinins, gibberellins and indole acetic acid. These beneficial bacteria also induce plant stress defense mechanisms and aid in pathogen suppression. Some free-living bacterial inoculants available on the market may provide biostimulant effects that justify application in their own right and outweigh the benefit of N fixation alone.
Research into biological N fixation in agricultural crops still has a long way to go, but it is clear that under the right circumstances, N-fixing bacteria can help growers produce healthy crops and offset N fertilizer requirements. Regulatory pressure to cut back on N use, as well as high fertilizer prices, motivates growers and consultants to look for new N management tools, and beneficial bacterial inoculants are worth serious consideration. Successful colonization and N fixation depend on field conditions and the degree of compatibility between microbial strain and crop type. Look for inoculants that have been tested on your crop and inquire about the bacteria’s tolerance to temperature, soil pH, moisture and ag chemicals. Further research and field trials are needed to improve consistent efficacy, but biological N fixation has the potential to become an important tool in crop production and environmental protection.
References
Aasfar A, Bargaz A, Yaakoubi K, Hilali A, Bennis I, Zeroual Y and Meftah Kadmiri I (2021) Nitrogen Fixing Azotobacter Species as Potential Soil Biological Enhancers for Crop Nutrition and Yield Stability. Front. Microbiol. 12:628379. doi: 10.3389/fmicb.2021.628379
Evers GW. Cool Season Legume Nitrogen Fixation. Texas A&M AgriLife Research and Extension Center at Overton. Retrieved 7/31/25. https://overton.tamu.edu/faculty-staff/gerald-wayne-evers/cool-season-legumes/nitrogen-fixation/
Guo K., Yang J., Yu N., Luo L., and Wang E. (2023). Biological nitrogen fixation in cereal crops: Progress, strategies, and perspectives. Plant Comm. 4, 100499.
Mahmud K, Makaju S, Ibrahim R, and Missaoui A. (2020). Current Progress in Nitrogen Fixing Plants and Microbiome Research. Plants. 9, 97. doi:10.3390/plants9010097
Rethwisch M. (2024) New/Recent Nitrogen Fixing Bacteria Products for Agriculture. University of California Agricultural and Natural Resources, Imperial County Agricultural Briefs. 27:1.
Science History Institute. (2025, July 29). Fritz Haber. Scientific Biographies, Science History Institute. https://www.sciencehistory.org/education/scientific-biographies/fritz-haber.
Yuja, S. 2023. Nitrogen-fixing organisms for sale? North Dakota State University Impact Stories. Jan 22, 2023. https://www.ndsu.edu/agriculture/ag-hub/impact-stories/nitrogen-fixing-organisms-sale
















 Invading California")

