

Grapes
Grapevine Red Botch Virus in Comparison to Grapevine Leafroll-Associated Virus
Grapevine Red Blotch associated Virus (GRBaV) is the causal viral agent responsible for Grapevine Red Blotch Disease. This…
Read ArticleArticle Archive
Grapevine Red Blotch associated Virus (GRBaV) is the causal viral agent responsible for Grapevine Red Blotch Disease. This…
Read Article
Applying fertilizers or plant nutrients to foliage has a long history, and there is an extensive number of…
Read Article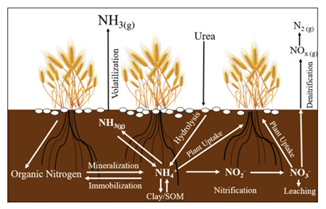
There is no doubt that almond producers have had to navigate numerous challenges over the past several years.…
Read Article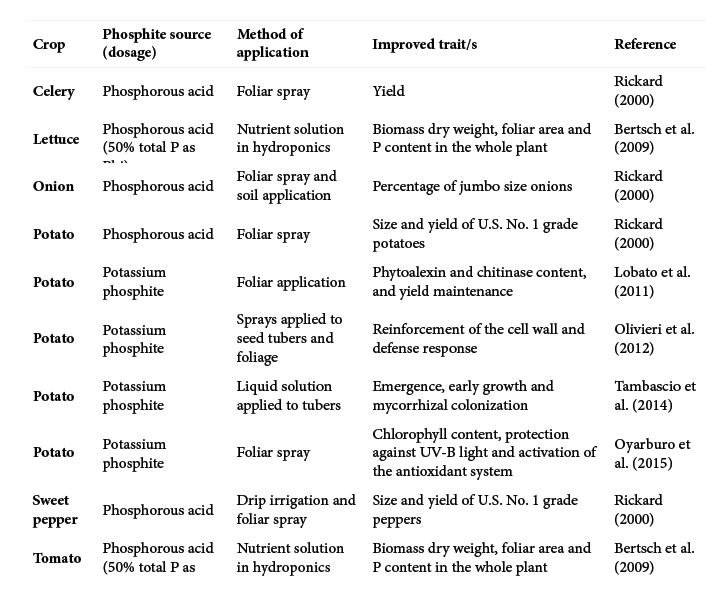
Phosphite has been a controversial topic for years. Its use and benefits are argued in hundreds of research…
Read Article
Most rice in California is established and grown in what is referred to as a water-seeded system. In…
Read Article
Walnuts are generally regarded as very sensitive to water stress. Severe stress and defoliation can occur when irrigation…
Read Article
Citrus mealybug infestations continue to increase in the San Joaquin Valley (SJV) making this species an emerging concern…
Read Article
Many of the pesticide application technologies used in specialty crop production today are based on axial fan air-blast…
Read Article
You will probably not surprise to see the grafting of fruit and nut tree crops and well understand…
Read Article
Whether your grapes are destined for wine, raisins, juice or the table, the job of the grapevine is…
Read Article